
Historical Aerial Photos (108,671 digital images)
The Water Resources Archives has a large collection of aerial photos. Below is a list of how our aerial photographs are organized, by broad geographic locations. The name in parentheses following the date and name of the area where the photo was taken refers to the collection to which the image belongs. The following collections are currently included in the Archives: Joseph Andrew Rowe (Rowe), Jim Imbiorski (Imbiorski), Natural Resources Conservation Service (NRCS), the Riverside County Flood Control and Water Conservation District (RCFCWCD), the San Bernardino County Flood Control District (SBCFCD), Lou Schnepp (Schnepp), San Bernardino County Archives (SBCA), and the United States Department of Agriculture via the San Bernardino County Flood Control District (USDA-SBCFCD). If you have aerial images to donate or would like to purchase high-resolution copies of images from the collection, please contact the Water Resources Archives at archives@csusb.edu.
Los Angeles County (252 Images)
Cities that are included in this section include Bel Air, Duarte, Garden Grove, Pomona, Walnut, Whittier and other cities and areas of Los Angeles County.
- 1928 Northside of San Gabriel Mountains - Flight C300 -- (Rowe) 36 images
- 1938 Whittier Narrows - Flight 38-16 -- (Rowe) 17 images
- 1940 Big Rock Creek - Flight AXJ307 -- (Rowe) 5 images
- 1940 Big Rock Creek - Flight AXJ308 -- (USDA-SBCFCD) 24 images
- 1945 Santa Clara River - Flight 9800 -- (Rowe) 17 images
- 1949 Bel Air Flight SPV -- (Rowe) 3 images
- 1952 Bel Air SFV1 -- (Rowe) 103 images
- 1955 San Gabriel River - Flight 9 -- (Rowe) 3 images
- 1955 San Gabriel River - Flight 10 -- (Rowe) 4 images
- 1982 Duarte -- (Imbiorski) 2 images
- 1991 Pomona - Flight 158-91 -- (Imbiorski) 3 images
- 1991 Walnut - Flight 139-91 -- (Imbiorski) 3 images
- 2000 Los Angeles County -- (Imbiorski) 9 images
- Garden Grove -- 6 images
Orange County (237 Images)
Cities that are included in this section include Laguna Beach, Santa Ana, and other cities that are located near the Santa Ana River.
- 1938 Lower Santa Ana River Flood Conditions - Flight 5029 -- (Rowe) 136 images
- 1946 Santa Ana River - Flight 311 -- (Rowe) 6 images
- 1952 Santa Ana Country Club -- (Rowe) 1 image
- 1955 Prado Dam-Santa Ana River - Flight 21677 -- (Rowe) 5 images
- 1962 Lower Santa Ana River area - Flight 24244 -- (Rowe) 20 images
- 1963-64 Prado Dam - Flight 5928 -- (Rowe) 9 images
- 1964 Santa Ana - Flight 5928 -- (Rowe) 31 images
- 1964 Lower Santa Ana River-Orange County -- (Rowe) 19 images
- 1991 Laguna Beach - Flight 146-91 -- (Imbiorski) 6 images
- Lower Santa Ana River area -- (Rowe) 3 images
- Prado Dam Santa Ana River-- Flight 5928 -- (Rowe) 1 image
Riverside County (75,036 Images)
-
Anza (516 images)
Anza is a semi-arid region located in south Riverside County in the Anza Valley. The small hamlet of Aquanga is also in this area. This area is mostly unincorporated areas of the high desert.
- 1958 Anza - RC-36-45 -- (RCFCWCD) 10 images
- 1961 Anza - RC-5-18 -- (RCFCWCD) 4 images
- 1961 Blythe - RC-34-37 -- (RCFCWCD) 4 images
- 1967 Burnt Valley - RC-1-38-1967 -- (RCFCWCD) 38 images
- 1967 Burnt Valley - RC-49-94-1967 -- (RCFCWCD) 45 images
- 1968 Anza - Horse Canyon - RC-1-25-1968 -- (RCFCWCD) 25 images
- 1970 Anza - RC-1-28-1970 -- (RCFCWCD) 28 images
- 1970 Anza - RC-9-27-1070 -- (RCFCWCD) 18 images
- 1970 Wilson Valley - RC-12-36-1970 -- (RCFCWCD) 21 images
- 1974 Long Canyon / West Wide Canyon -RC-1-82-1974 -- (RCFCWCD) 81 images
- 1975 Anza Disposal -RC-1-3-1975 -- (RCFCWCD) 3 images
- 1976 Tripp Flats Area - RC-Anza-1-26-1976 -- (RCFCWCD) 26 images
- 1977 Anza Disposal - RC-Anza-1-3-1977 -- (RCFCWCD) 3 images
- 1978 Anza Disposal -RC-Anza-1-3-1978 -- (RCFCWCD) 3 images
- 1979 Anza Disposal - RC-Anza-2-3-5-1979 -- (RCFCWCD) 3 images
- 1983 Anza - RC-Anza-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Anza Sanitary Landfill - RC-Anza-1-3-1983 -- (RCFCWCD) 3 images
- 1983 Anza Sanitary Landfill - RC-Anza-2-4-1983 -- (RCFCWCD) 3 images
- 1985 Anza Sanitary Landfill - RC-1-3-1985 -- (RCFCWCD) 3 images
- 1985 Long Canyon Mapping - RC-1-8-1985 -- (RCFCWCD) 8 images
- 1986 Anza Sanitary Landfill - RC-Anza-1-3-1986 -- (RCFCWCD) 3 images
- 1987 Long Canyon Mapping - RC-1-24-1987 -- (RCFCWCD) 24 images
- 1987 Nicholls Warm Spring - RC-1-12-1987 -- (RCFCWCD) 12 images
- 1987 Nicholls Warm Spring - RC-13-27-1987 -- (RCFCWCD) 15 images
- 1989 Anza Sanitary Landfill - RC-1-3-1989 -- (RCFCWCD) 3 images
- 1992 Anza Sanitary Landfill - RC-Anza-1-3-1992 -- (RCFCWCD) 3 images
- 1993 Anza Sanitary Landfill - RC-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Wilson Creek - RC-1-9-1993 -- (RCFCWCD) 9 images
- 1994 Anza Sanitary Landfill - RC-Anza2-1-3-1994 -- (RCFCWCD) 3 images
- 1994 Anza Sanitary Landfill - RC-Anza-1-3-1994 -- (RCFCWCD) 3 images
- 1997 Anza Sanitary Landfill - RC-Anza2-1-3-1997 -- (RCFCWCD) 3 images
- 1997 Anza Sanitary Landfill Spot - RC-Anza-A-1997 -- (RCFCWCD) 1 image
- 1998 Anza Sanitary Landfill - RC-1-3-1998 -- (RCFCWCD) 3 images
- 1998 Anza Sanitary Landfill Spot - RC-1998 -- (RCFCWCD) 1 image
- 1999 Anza Sanitary Landfill - RC-Anza-1-3-1999 -- (RCFCWCD) 3 images
- 2000 Anza Sanitary Landfill - RC-Anza-1-3-2000 -- (RCFCWCD) 3 images
- 2002 Anza Sanitary Landfill - RC-1-5-2002 -- (RCFCWCD) 5 images
- 2005 Anza Sanitary Landfill - RC-Anza-1-3-2005 -- (RCFCWCD) 3 images
- 2008 Anza Ortho - RC-1-65-2008 -- (RCFCWCD) 65 images
- 2010 Long Canyon - RC-1-20-2010 -- (RCFCWCD) 20 images
Corona (6,264 images)
The cities of Corona, Chino Hills, Yorba Linda, Jurupa Valley, and Norco are located in the western end of the County of Riverside.
- 1931 Riverside-Corona - RC-1-373-1931 -- (RCFCWCD) 217 images
- 1931 Riverside-Corona - RC-Corona-1-35-52-1931 -- (RCFCWCD) 8 images
- 1931 Riverside-Corona - RC-Corona-2-1-3-1931 -- (RCFCWCD) 3 images
- 1931 Riverside-Corona - RC-Corona-10-1-2-1931 -- (RCFCWCD) 2 images
- 1931 Riverside-Corona - RC-Corona-11-2-1931 -- (RCFCWCD) 1 image
- 1939 Temescal Canyon/Coldwater Canyon - Flight 5928-1-11-1939 --( Rowe) 10 images
- 1947 Lake Matthews - RC-9-1947 -- (RCFCWCD) 1 image
- 1948 Corona to Murrieta North of Lake ELsinore - RC-61-322-1948 -- (RCFCWCD) 177 images
- 1948 Corona to Murrieta South of Lake Elsinore - RC-67-360-1948 -- (RCFCWCD) 239 images
- 1948 Northwest County Areas - RC-45-76-1948 -- (RCFCWCD) 12 images
- 1948 Prado Basin- RC-1-13-1948 -- (RCFCWCD) 12 images
- 1949 Corona Book 6A - RC-AXM-F-3F-2-205-1949 -- (RCFCWCD) 35 images
- 1949 Corona Book 6A - RC-AXM-F-4F-30-190-1949 -- (RCFCWCD) 32 images
- 1952 Santa Ana River & Prado Basin - RC-1-1-110-1952 -- (RCFCWCD) 104 images
- 1952 Santa Ana River & Prado Basin - RC-2-1-15-1952-- (RCFCWCD) 15 images
- 1956 Mira Loma/Norco - RC-3-1-32 -- (RCFCWCD) 33 images
- 1957 Corona - RC-5-37-40 -- (RCFCWCD) 4 images
- 1958 Corona - RC-5-11-22 -- (RCFCWCD) 12 images
- 1958 Corona - RC-1-9 -- (RCFCWCD) 12 images
- 1958 El Cerrito - RC-14-36 -- (RCFCWCD) 20 images
- 1959 Norco - RC-1-13 -- (RCFCWCD) 13 images
- 1959 Norco - RC-5-8 -- (RCFCWCD) 4 images
- 1960 Corona - RC-21-23-1960 -- (RCFCWCD) 3 images
- 1960 Norco/Santa Ana River - RC-35-49-1961 -- (RCFCWCD) 15 images
- 1960 North Norco Channel - RC-1-20-1960 -- (RCFCWCD) 20 images
- 1960 North Norco Channel Convergence - RC-24-66-1960 -- (RCFCWCD) 40 images
- 1961 Cajelco Road - RC-1-1-1-56 -- (RCFCWCD) 56 images
-
1961 Corona - RC-23-71 -- (RCFCWCD) 49 images
- 1961 Corona - RC-60-62 -- (RCFCWCD) 3 images
- 1961 Lake Matthews - RC-63-69 -- (RCFCWCD) 7 images
- 1961 South Norco Channel - RC-44-82 -- (RCFCWCD) 39 images
- 1963 Corona - RC-HA-UC-3-68-1963 -- (RCFCWCD) 45 images
- 1963 Corona-Skyline - RC-1-12-1963 -- (RCFCWCD) 39 images
- 1963 North Norco Channel - RC-41-81-1963 -- (RCFCWCD) 41 images
- 1963 Temescal Canyon - RC-148-172-1963 -- (RCFCWCD) 25 images
- 1965 Cajalco - RC-50-98-1965 -- (RCFCWCD) 50 images
- 1965 Prado Dam - RC-41-82-1965 -- (RCFCWCD) 44 images
- 1967 Lincoln St. Drain - RC-7-21-1967 -- (RCFCWCD) 15 images
- 1968 Norco Revision - RC-20-54-1968 -- (RCFCWCD) 35 images
- 1969 Oak Street - Corona - RC-1-22-1969 -- (RCFCWCD) 22 images
- 1969 Oak Street Channel - RC-9-20-1969 -- (RCFCWCD) 11 images
- 1969 Temescal -RC-77-88-1969 -- (RCFCWCD) 12 images
- 1969 Temescal Creek -RC-12-29-1969 -- (RCFCWCD) 18 images
- 1970 Temescal Canyon - RC-1-12-1970 -- (RCFCWCD) 7 images
- 1971 Cajalco - RC-1-13-1971 -- (RCFCWCD) 13 images
- 1971 Corona Dumpsite - RC-Dump-1-3-1971 -- (RCFCWCD) 3 images
- 1971 Corona Main Street Convergents - RC-5A-15B-1971 -- (RCFCWCD) 21 images
- 1971 Corona Main Street Wash - RC-16-48-1971 -- (RCFCWCD) 33 images
- 1971 Corona Raceway - RC-Raceway-1-1971 -- (RCFCWCD) 1 image
- 1972 Corona Revision - RC-28-74-1972 -- (RCFCWCD) 42 images
- 1972 Harley John Road - RC-37-52-1972 -- (RCFCWCD) 16 images
- 1972 Lake Matthews West - RC-11-30-1972 -- (RCFCWCD) 20 images
- 1972 Maley Canyon - RC-1-7-1972 -- (RCFCWCD) 7 images
- 1972 Maley Canyon High Obliques - RC-9-1972 -- (RCFCWCD) 1 image
- 1973 Maybe Canyon - RC-106-109-1973 -- (RCFCWCD) 2 images
- 1973 Maybe Canyon - RC-110-111-1973 -- (RCFCWCD) 2 images
- 1973 Maybe Canyon - RC-112-121-1973 -- (RCFCWCD) 2 images
- 1973 North Norco Channel -RC-1-50-1973 -- (RCFCWCD) 50 images
- 1974 Corona Dumpsite - RC-3-5-1974 -- (RCFCWCD) 3 images
- 1974 Corona Dumpsite Convergence - RC-Corona-1-2-1974 -- (RCFCWCD) 2 images
- 1974 Mockingbird Canyon - RC-1-31-1974 -- (RCFCWCD) 31 images
- 1974 Oak Street Corona - RC-15-30-1974 -- (RCFCWCD) 16 images
- 1974 Oak Street - Corona Convergence - RC-1-14-1974 -- (RCFCWCD) 14 images
- 1975 Lake Evans - RC-10-11-1975 -- (RCFCWCD) 2 images
- 1975 Main Street Corona - RC-1-4-1975 -- (RCFCWCD) 4 images
- 1975 Main Street Corona High Obliques -RC-5-6-1975 -- (RCFCWCD) 2 images
- 1976 Corona Disposal - RC-Corona-3-5-1976 -- (RCFCWCD) 3 images
- 1976 North Norco Channel - RC-1-1-5-45-1976 -- (RCFCWCD) 45 images
- 1977 Corona Burn - RC-Corona-1-6-1977 -- (RCFCWCD) 6 images
- 1977 Corona Burn Color Prints - RC-1-126-1977 -- (RCFCWCD) 125 images
- 1977 Corona Burn Obliques - RC-7-23-1977 -- (RCFCWCD) 17 images
- 1977 Corona Disposal - RC-Corona-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Mead Valley Disposal - RC-Mead-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Mockingbird Canyon - RC-1-41-1977 -- (RCFCWCD) 41 images
- 1978 Corona Disposal - RC-Corona-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Corona Flood - RC-1-40-1978 -- (RCFCWCD) 38 images
- 1978 Mabey Canyon - RC-1-2-1978 -- (RCFCWCD) 2 images
- 1978 Main Street Basin - RC-5-9-1978 -- (RCFCWCD) 5 images
- 1978 Mockingbird Canyon - RC-1-11-1978 -- (RCFCWCD) 11 images
- 1978 Oak Street Basin - RC-1-21-1978 -- (RCFCWCD) 23 images
- 1978 Section 6, T4S, R4W - RC-Section-6-1-3-1978 -- (RCFCWCD) 3 images
- 1979 Corona - RC-2-3-7-1979 -- (RCFCWCD) 5 images
- 1979 Corona Drain Extension - RC-1-16-1979 -- (RCFCWCD) 15 images
- 1979 Mabey Canyon - RC-Mabey-1-2-1979 -- (RCFCWCD) 2 images
- 1979 Main Street Basin - RC-5-9-1979 -- (RCFCWCD) 2 images
- 1979 Mockingbird Canyon - RC-Mockingbird-1-55-1979 -- (RCFCWCD) 55 images
- 1979 North Norco - RC-1-21-1979 -- (RCFCWCD) 19 images
- 1979 Oak Street Basin - RC-1-3-1979 -- (RCFCWCD) 3 images
- 1979 Oak Street Basin Obliques - RC-4-6-1979 -- (RCFCWCD) 3 images
- 1980 City of Corona - RC-1-35-1980 -- (RCFCWCD) 35 images
- 1980 City of Corona - RC-49-72-1980 -- (RCFCWCD) 24 images
- 1980 City of Corona - RC-73-80-1980 -- (RCFCWCD) 8 images
- 1980 City of Corona - RC-81-90-1980 -- (RCFCWCD) 10 images
- 1980 City of Corona - RC-91-104-1980 -- (RCFCWCD) 14 images
- 1980 City of Corona Ortho - RC-36-48-1980 -- (RCFCWCD) 13 images
- 1980 Cold Water Deep Canyon to Archibald - Flight 77-80 -- (Imbiorski) 11 images
- 1980 Corona Sand and Glass Company - RC-1-2-1980 -- (RCFCWCD) 2 images
- 1980 Corona Sanitary Drain - RC-1-2-1980 -- (RCFCWCD) 2 images
- 1980 Corona Sanitary Drain Ortho - RC-3-5-1980 -- (RCFCWCD) 3 images
- 1980 Mead Valley - RC-15-97-1980 -- (RCFCWCD) 83 images
- 1980 MeadValley - RC-Mead-1-14-1980 -- (RCFCWCD) 14 images
- 1980 North Norco Channel - RC-1-11-1980 -- (RCFCWCD) 11 images
- 1980 Temescal Creek Flood Damage - RC-1-22-1980 -- (RCFCWCD) 22 images
- 1980 Temescal Wash - RC-3-8-1980 -- (RCFCWCD) 6 images
- 1980 Temescal Wash - RC-1-97-1980 -- (RCFCWCD) 97 images
- 1981 City of Corona Reflight - RC-1-5-1981 -- (RCFCWCD) 5 images
- 1981 North Norco - RC-1-11-1981 -- (RCFCWCD) 11 images
- 1981 North Norco Channel - RC-2-15-3-24-1981 -- (RCFCWCD) 2 images
- 1981 Temescal Creek - RC-1-32-1981 -- (RCFCWCD) 33 images
- 1982 Corona Sanitary Landfill - RC-8-10-1982 -- (RCFCWCD) 3 images
- 1982 Mead Valley - RC-1-21-1982 -- (RCFCWCD) 21 images
- 1982 Mead Valley Ortho - RC-22-83-1982 -- (RCFCWCD) 62 images
- 1982 North Norco Channel - RC-Norco-1-18-1982 -- (RCFCWCD) 18 images
- 1982 Temescal - RC-1-6-1982 -- (RCFCWCD) 6 images
- 1982 Temescal - RC-11-32-1982 -- (RCFCWCD) 22 images
- 1983 Columbia Basin - RC-Columbia-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Corona Sanitary Landfill - RC-Corona-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Corona Sanitary Landfill - RC-1-A-10-1983 -- (RCFCWCD) 8 images
- 1983 Corona Sanitary Landfill - RC-3-7-1983 -- (RCFCWCD) 5 images
- 1983 Glen Ivy - RC-1-1983 -- (RCFCWCD) 1 image
- 1983 Home Gardens - RC-1-3-1983 -- (RCFCWCD) 3 images
- 1983 Mabey Canyon - RC-Mabey-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Main Street - RC-Main-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Mead Valley Sanitary Landfill - RC-Mead-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Mockingbird Canyon - RC-1-5-1983 -- (RCFCWCD) 5 images
- 1983 South Norco Channel - RC-4-10-1983 -- (RCFCWCD) 7 images
- 1983 South Norco Channel - RC-Norco-1-22-1983 -- (RCFCWCD) 22 images
- 1983 Oak Street - RC-Oak-1-4-1983 -- (RCFCWCD) 4 images
- 1984 Corona Sanitary Landfill - RC-Corona-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Corona Sanitary Landfill - RC-Corona-1-5-1984 -- (RCFCWCD) 5 images
- 1984 Corona Sanitary Landfill - RC-Corona-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Corona Sanitary Landfill - RC-Corona-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Corona Sanitary Landfill 6/84 Extension - RC-Corona-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Mabey Canyon Basin - RC-Mabey-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Main Street Basin - RC-Main-1-5-1984 -- (RCFCWCD) 5 images
- 1984 Mockingbird Sanitary Landfill - RC-Mockingbird-1-4-1984 -- (RCFCWCD) 4 images
- 1984 North Norco Channel - RC-1-9-1984 -- (RCFCWCD) 9 images
- 1984 North Norco Line NA - RC-1-56-1984 -- (RCFCWCD) 56 images
- 1984 Oak Street Basin - RC-Oak-1-9-1984 -- (RCFCWCD) 5 images
- 1984 Prado Basin - RC-Prado-1-7-1984 -- (RCFCWCD) 7 images
- 1984 Temescal Creek Channel - RC-1-18-1984 -- (RCFCWCD) 15 images
- 1985 El Cerrito Channel - RC-1-4-1985 -- (RCFCWCD) 4 images
- 1985 Mabey Canyon Basin - RC-Mabey-1-4-1985 -- (RCFCWCD) 4 images
- 1985 Main Street Basin - RC-Main-1-5-1985 -- (RCFCWCD) 5 images
- 1985 Mockingbird - RC-Mockingbird-1-4-1985 -- (RCFCWCD) 4 images
- 1985 North Main Street Channel - RC-North-1-4-1985 -- (RCFCWCD) 4 images
- 1987 Mabey Canyon Basin - RC-Mabey-1-5-1987 -- (RCFCWCD) 5 images
- 1987 North Main Street Channel - RC-Main-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Main Street Basin - RC-Main2-1-5-1987 -- (RCFCWCD) 4 images
- 1987 Mockingbird Canyon - RC-Mockingbird-1-3-1987 -- (RCFCWCD) 3 images
- 1987 Norco Mapping - RC-1-22-1987 -- (RCFCWCD) 22 images
- 1987 Norco Ortho - RC-1-8-1987 -- (RCFCWCD) 8 images
- 1987 Oak Street Basin - RC-Oak-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Temescal Channel - RC-Temescal-1-4-1987 -- (RCFCWCD) 4 images
- 1988 Mabey Canyon - RC-Mabey-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Main Street - RC-Main-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Mockingbird Canyon - RC-Mockingbird-1-3-1988 -- (RCFCWCD) 3 images
- 1988 Oak Street - RC-Oak-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Prado Spot - RC-Prado-1-1988 -- (RCFCWCD) 1 image
- 1988 Temescal Canyon - RC-1-30-1988 -- (RCFCWCD) 30 images
- 1989 Mabey Canyon - RC-Mabey-1-3-1989 -- (RCFCWCD) 3 images
- 1989 Main Street Basin - RC-Main-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Mockingbird Canyon - RC-Mockingbird-1-3-1989 -- (RCFCWCD) 3 images
- 1989 Oak Street Basin - RC-Oak-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Traffic Signals (Corona) - RC-9-35-1989 -- (RCFCWCD) 24 images
- 1990 Mabey Canyon Basin - RC-Mabey-1-3-1990 -- (RCFCWCD) 3 images
- 1990 Main Street Basin - RC-1-4-1990 -- (RCFCWCD) 4 images
- 1990 Mead Valley Sanitary Landfill -RC-1-10-1990 -- (RCFCWCD) 10 images
- 1990 Oak Street - RC-1-19-1990 -- (RCFCWCD) 19 images
- 1990 Oak Street Basin - RC-Oak-1-4-1990 -- (RCFCWCD) 4 images
- 1990 Wood Road - RC-1-9-1990 -- (RCFCWCD) 9 images
- 1991 Corona Sanitary Landfill - RC-1-5-1991 -- (RCFCWCD) 5 images
- 1991 Mabey Canyon Basin - RC-Mabey-1-3-1991 -- (RCFCWCD) 3 images
- 1991 Main Street Basin - RC-Main-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Main Street Channel - RC-1-16-1991 -- (RCFCWCD) 16 images
- 1991 Mockingbird Canyon - RC-Mockingbird-1-10-1991 -- (RCFCWCD) 10 images
- 1991 Oak Street Basin - RC-Oak-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Oak Street Channel - RC-Oak-1-16-1991 -- (RCFCWCD) 16 images
- 1991 Route 91 & Gran River - RC-1-19-1991 -- (RCFCWCD) 19 images
- 1991 South Corona Channel - RC-South-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Temescal Basin - RC-Temescal-1-5-1991 -- (RCFCWCD) 5 images
- 1991 Temescal Channel - RC-Temescal2-1-5-1991 -- (RCFCWCD) 5 images
- 1991 Temescal Creek Wash R/W - RC-1-2-1991 -- (RCFCWCD) 2 images
- 1992 Mabey Canyon Basin - RC-Mabey-1-3-1992 -- (RCFCWCD) 3 images
- 1992 Main Street Basin - RC-Main-1-5-1992 -- (RCFCWCD) 5 images
- 1992 Oak Street Basin - RC-Oak-1-5-1992 -- (RCFCWCD) 5 images
- 1993 Mabey Canyon - RC-Mabey-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Main Street Basin - RC-Main-1-4-1993 -- (RCFCWCD) 4 images
- 1993 Mockingbird Canyon - RC-Mockingbird-1-7-1993 -- (RCFCWCD) 7 images
- 1993 Oak Street Basin - RC-Oak-1-4-1993 -- (RCFCWCD) 4 images
- 1993 South Norco Line SA - RC-Norco-1-2-1993 -- (RCFCWCD) 2 images
- 1993 Temescal Canyon - RC-1-91-1993 -- (RCFCWCD) 91 images
- 1994 Cajalco Road - RC-Cajalco-1-25-1994 -- (RCFCWCD) 25 images
- 1994 Corona Spot - RC-Corona-A-1994 -- (RCFCWCD) 1 image
- 1994 Corona Sanitary Landfill - RC-Corona2-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Mabey Canyon - RC-Mabey-1-3-1994 -- (RCFCWCD) 4 images
- 1994 Main Street Basin - RC-Main-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Norco Bluff Study - RC-1-15-1994 -- (RCFCWCD) 15 images
- 1994 Norco Bluff Study - RC-1-21-1994 -- (RCFCWCD) 21 images
- 1994 Norco Bluff Study - RC-1-22-1994 -- (RCFCWCD) 22 images
- 1995 Norco Norco Master Drainage Plan, Line NA-5 - RC-1-4-1995 -- (RCFCWCD) 4 images
- 1995 Norco Bluff Study - RC-22-36-1995 -- (RCFCWCD) 15 images
- 1995 Norco Bluff Study - RC-23-37-1995 -- (RCFCWCD) 15 images
- 1994 Oak Street Channel - RC-1-30-1994 -- (RCFCWCD) 30 images
- 1994 South Norco Channel - RC-1A-4B-1994 -- (RCFCWCD) 9 images
- 1995 Corona Sanitary Landfill - RC-Corona2-1-4-1995 -- (RCFCWCD) 4 images
- 1995 Corona Sanitary Landfill Spot - RC-Corona-A-1995 -- (RCFCWCD) 1 image
- 1995 Mabey Canyon - RC-Mabey-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Norco Corona Ortho - RC-1-83-1995 -- (RCFCWCD) 83 images
- 1995 Norco Corona Ortho - RC-84-130-1995 -- (RCFCWCD) 47 images
- 1995 Norco Bluff Study - RC-22-36-1995 -- (RCFCWCD) 15 images
- 1995 Norco Bluff Study - RC-23-37-1995 -- (RCFCWCD) 15 images
- 1995 Temescal Canyon - RC-1-09-1995 -- (RCFCWCD) 9 images
- 1996 Corona Sanitary Landfill - RC-Corona-1-4-1996 -- (RCFCWCD) 4 images
- 1996 East Matthews Ortho - RC-East-1-75-1996 -- (RCFCWCD) 80 images
- 1996 Estelle Street Storm Drain - RC-Estelle-1-3-1996 -- (RCFCWCD) 3 images
- 1996 Mabey Canyon - RC-Mabey-1-3-1996 -- (RCFCWCD) 3 images
- 1996 Main Street Basin - RC-Main-1-6-1996 -- (RCFCWCD) 6 images
- 1996 McBride Canyon - RC-McBride-1-13-1996 -- (RCFCWCD) 13 images
- 1996 Mockingbird Canyon - RC-Mockingbird-1-6-1996 -- (RCFCWCD) 6 images
- 1996 Oak Street Dam - RC-Oak-1-3-1996 -- (RCFCWCD) 3 images
- 1997 Brown Canyon Channel - RC-1-11-1997 -- (RCFCWCD) 11 images
- 1997 Cajalco Lateral - RC-Cajalco-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Corona Compton Avenue - RC-Corona-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Corona Drain Line 7-A - RC-Corona-1-18-1997 -- (RCFCWCD) 18 images
- 1997 Mabey Canyon Basin - RC-Mabey-1-3-1997 -- (RCFCWCD) 3 images
- 1997 Main Street Basin - RC-Main-1-6-1997 -- (RCFCWCD) 6 images
- 1997 McBride Canyon Channel - RC-1-13-1997 -- (RCFCWCD) 13 images
- 1997 Mockingbird Canyon - RC-Mockingbird-1-12-1997 -- (RCFCWCD) 12 images
- 1997 Norco Bluffs - RC-1-26-1997 -- (RCFCWCD) 26 images
- 1997 Norco Master Drainage Plan - RC-Norco-1-7-1997 -- (RCFCWCD) 7 images
- 1997 Oak Street Basin - RC-1-43-1997 -- (RCFCWCD) 43 images
- 1997 Temescal - RC-Temescal-1-6-1997 -- (RCFCWCD) 6 images
- 1997 Upper Temescal - RC-Upper-1-5-1997 -- (RCFCWCD) 4 images
- 1998 Brown Canyon Channel - RC-Brown-1-9-1998 -- (RCFCWCD) 9 images
- 1998 Eastvale - RC-1-117-1998 -- (RCFCWCD) 117 images
- 1998 Mabey Canyon Basin - RC-Mabey-1-3-1998 -- (RCFCWCD) 3 images
- 1998 Main Street Basin - RC-Main-1-5-1998 -- (RCFCWCD) 5 images
- 1998 Mockingbird Canyon - RC-1-13-1998 -- (RCFCWCD) 13 images
- 1998 Oak Street Dam and Channel - RC-1-41-1998 -- (RCFCWCD) 41 images
- 1998 Temescal Canyon - RC-1-15-1998 -- (RCFCWCD) 15 images
- 1999 Brown Canyon - RC-Brown-1-12-1999 -- (RCFCWCD) 12 images
- 1999 Galvan / Smith Road - RC-1-8-1999 -- (RCFCWCD) 8 images
- 1999 Mabey Canyon - RC-Mabey-1-3-1999 -- (RCFCWCD) 3 images
- 1999 Main Street Dam - RC-Main-1-6-1999 -- (RCFCWCD) 5 images
- 1999 McBride Canyon Channel - RC-1-14-1999 -- (RCFCWCD) 14 images
- 1999 Mockingbird Canyon - RC-Mockingbird-1-12-1999 -- (RCFCWCD) 12 images
- 1999 Oak Street Channel - RC-1-43-1999 -- (RCFCWCD) 43 images
- 1999 Oak Street Dam - RC-Oak-1-3-1999 -- (RCFCWCD 3 images
- 1999 Temescal Creek Dam - RC-Temescal-1-15-1999 -- (RCFCWCD) 15 images
- 2000 Corona Sanitary Landfill - RC-Corona2-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Brown Canyon - RC-1-1-2-7-2000 -- (RCFCWCD) 10 images
- 2000 Corona Sanitary Landfill Spot - RC-Corona-A-2000 -- (RCFCWCD) 1 image
- 2000 Mabey Canyon Basin - RC-Mabey-1-3-2000 -- (RCFCWCD) 3 images
- 2000 Main Street Dam - RC-Main-1-5-2000 -- (RCFCWCD) 5 images
- 2000 McBride Channel - RC-1-1-4-3-2000 -- (RCFCWCD) 13 images
- 2000 Mockingbird Canyon - RC-1-1-2-6-2000 -- (RCFCWCD) 12 images
- 2000 Oak Street Channel - RC-1-37-2000 -- (RCFCWCD) 37 images
- 2000 Oak Street Dam - RC-Oak-1-3-2000 -- (RCFCWCD) 3 images
- 2000 Temescal Creek - RC-1-1-3-4-2000 -- (RCFCWCD) 15 images
- 2001 Alberhill Ortho - RC-1-70-2001 -- (RCFCWCD) 69 images
- 2001 Brown Canyon Channel - RC-Brown-1-10-2001 -- (RCFCWCD) 10 images
- 2001 Brown Canyon McBride - RC-1-13-2001 -- (RCFCWCD) 13 images
- 2001 I-15 Ortho - RC-1-47-2001 -- (RCFCWCD) 47 images
- 2001 Mabey Canyopn Dam / Basin - RC-Mabey-1-3-2001 -- (RCFCWCD) 3 images
- 2001 Main Street Dam - RC-Main-1-5-2001 -- (RCFCWCD) 5 images
- 2001 Mockingbird Canyon - RC-Mockingbird-1-14-2001 -- (RCFCWCD) 14 images
- 2001 Oak Street Dam & Channel - RC-1-42-2001 -- (RCFCWCD) 42 images
- 2001 Special Project Ortho - RC-1-23-2001 -- (RCFCWCD) 23 images
- 2001 Temescal Creek Channel - RC-Temescal-1-16-2001 -- (RCFCWCD) 16 images
- 2002 Corona Drain Line 46 extension - RC-Corona-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Mabey Canyon Dam - RC-Mabey-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Main Street Dam - RC-Main-1-4-2002 -- (RCFCWCD) 4 images
- 2002 Mockingbird Canyon - RC-1-9-2002 -- (RCFCWCD) 9 images
- 2002 Temescal Basin Cajalco Lateral - RC-1-1-2-4-2002 -- (RCFCWCD) 8 images
- 2002 West Norco Basin - RC-West-1-2-2002 -- (RCFCWCD) 2 images
- 2003 Corona Drains Line 46 Extension II - RC-Corona-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Eastvale Basin - RC-Eastvale-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Eastvale Basin - RC-Eastvale2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Eastvale Ortho - RC-1-50-2003 -- (RCFCWCD) 50 images
- 2003 Mabey Canyon Dam - RC-Mabey-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Main Strret Dam - RC-Main-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Oak Street Dam - RC-Oak2-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Oak Street Dam - RC-Oak-1-6-2003 -- (RCFCWCD) 6 images
- 2003 River Ranch Organics - RC-River-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Temescal Channel Cota Street -RC-Temescal-1-7-2003 -- (RCFCWCD) 7 images
- 2004 Eastvale Basin - RC-Eastvale-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Lamb Canyon Sanitary Landfill - RC-1-21-2004 -- (RCFCWCD) 21 images
- 2004 Mabey Canyon Dam - RC-Mabey-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Main Street Dam - RC-Main-1-4-2004 -- (RCFCWCD) 4 images
- 2004 Mockingbird Canyon - RC-1-9-2004 -- (RCFCWCD) 9 images
- 2004 Mockingbird Canyon - RC-1-10-2004 -- (RCFCWCD) 10 images
- 2004 Oak Street Dam - RC-1-6-2004 -- (RCFCWCD) 6 images
- 2004 Temescal Canyon - RC-1-7-2004 -- (RCFCWCD) 7 images
- 2004 West Norco Basin - RC-West-1-2-2004 -- (RCFCWCD) 2 images
- 2005 Agriscape - RC-Agriscape-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Agriscape 2 - RC-Agriscape2-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Agriscape 3 - RC-Agriscape3-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Agriscape 4 - RC-Agriscape4-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Corona Sanitary Landfill -RC-Corona2-1-4-2005 -- (RCFCWCD) 4 images
- 2005 Corona Sanitary Landfill Spot - RC-Corona-A-2005 -- (RCFCWCD) 1 image
- 2005 Eastvale Basin - RC-Eastvale-7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Mabey Canyon Dam - RC-Mabey7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Main Street Dam - RC-Main7-1-4-2005 -- (RCFCWCD) 4 images
- 2005 Mockingbird Canton - RC-Mockingbird7-1-9-2005 -- (RCFCWCD) 4 images
- 2005 Norco Bluffs - RC-Norco3-1-7-2005 -- (RCFCWCD) 7 images
- 2005 Norco Main Drain Pipe Line - RC-Norco2-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Norco Main Drain Pipe NA-3 - RC-Norco-1-4-2005 -- (RCFCWCD) 4 images
- 2005 South Norco Channel - RC-Norco7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Oak Street Dam - RC-Oak7-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Santa Ana River Prado Dam - RC-1-14-7-2005 -- (RCFCWCD) 32 images
- 2005 South Norco Channel A-SB Stage 3 - RC-Norco-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Southern California Recycling - RC-So-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Temescal Basin and Cajalco Lateral - RC-Temescal7-1-8-2005 -- (RCFCWCD) 8 images
- 2005 Temescal Channel/Cota Street - RC-Temescal8-1-7-2005 -- (RCFCWCD) 7 images
- 2005 West Norco Basin - RC-Norco7-1-2-2005 -- (RCFCWCD) 2 images
- 2006 Brown Canyon - RC-Brown-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Eastvale Basin - RC-Eastvale-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Hunt Canyon - RC-Hunt-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Mabey Canyon Dam - RC-Mabey-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Main Street Dam - RC-Main-1-4-2006 -- (RCFCWCD) 4 images
- 2006 Matthews Ortho - RC-1-63-2006 -- (RCFCWCD) 63 images
- 2006 Mockingbird Canyon - RC-Mockingbird-1-9-2006 -- (RCFCWCD) 9 images
- 2006 North Norco Channel Stage 9 - RC-Norco6-1-9-2006 -- (RCFCWCD) 9 images
- 2006 San Sevaine Channel - RC-Sevaine-1-7-2006 -- (RCFCWCD) 7 images
- 2006 South Norco Channel - RC-South-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Temescal Basin - RC-Temescal2-1-8-2006 -- (RCFCWCD) 2 images
- 2006 Temescal Channel - RC-Temescal-1-7-2006 -- (RCFCWCD) 7 images
- 2006 West Norco Basin - RC-West-1-2-2006 -- (RCFCWCD) 2 images
- 2007 Brown Canyon Debris Basin - RC-1-2007 -- (RCFCWCD) 1 image
- 2007 Eastvale Basin - RC-Eastvale-1-2-2007 -- (RCFCWCD) 2 images
- 2007 El Cerrito Ortho - RC-1-61-2007 -- (RCFCWCD) 61 images
- 2007 El Cerrito Ortho Spot - RC-El-Spot-2007 -- (RCFCWCD) 1 image
- 2007 Hunt Canyon Debris Basin - RC-Hunt-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Mabey Canyon Dam - RC-Mabey-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Main Street Dam - RC-Main2-1-4-2007 -- (RCFCWCD) 18 images
- 2007 Main Street Dam - RC-1-4-2007 -- (RCFCWCD) 4 images
- 2007 Mockingbird Canyon - RC-Mockingbird-1-9-2007 -- (RCFCWCD) 9 images
- 2007 Norco Channel Stage 9 - RC-Norco2-1-6-2007 -- (RCFCWCD) 6 images
- 2007 Oak Street Dam - RC-Oak2-1-6-2007 -- (RCFCWCD) 6 images
- 2007 Oak Street Dam Debris Basin - RC-1-6-2007 -- (RCFCWCD) 6 images
- 2007 Ontario Avenue - RC-1-18-2007 -- (RCFCWCD) 18 images
- 2007 South Norco Channel - RC-South-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Temescal Channel - RC-Temescal-1-7-2007 -- (RCFCWCD) 7 images
- 2007 Temescal Basin and Cajalco Lateral - RC-Temescal2-1-8-2007 -- (RCFCWCD) 8 images
- 2007 West Norco Basin - RC-West-1-2-2007 -- (RCFCWCD) 2 images
- 2008 Mabey Canyon - RC-Mabey-1-3-2008 -- (RCFCWCD) 3 images
- 2008 Main Street Drain - RC-Main-1-4-2008 -- (RCFCWCD) 4 images
- 2009 Brown Canyon Debris Basin - RC-Brown-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Brown Canyon Debris Basin - RC-Brown7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Cleveland Basin - RC-Cleveland-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Eastvale Basin - RC-Eastvale-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Eastvale Basin - RC-Eastvale7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Hunt Canyon Debris Basin - RC-Hunt-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Hunt Canyon Debris Basin - RC-Hunt7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Mabey Canyon Dam - RC-Mabey7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Mabey Canyon Debris Basin - RC-Mabey-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Main Street Dam - RC-Main-1-5-2009 -- (RCFCWCD) 5 images
- 2009 Main Street Dam - RC-Main7-1-4-2009 -- (RCFCWCD) 4 images
- 2009 Mockingbird Canyon - RC-Mockingbird-1-9-2009 -- (RCFCWCD) 9 images
- 2009 Mockingbird Canyon - RC-1-9-2009 -- (RCFCWCD) 9 images
- 2009 Norco-Corona Ortho (Part 1) - RC-1-252-2009 -- (RCFCWCD) 252 images
- 2009 Norco-Corona Ortho (Part 2) - RC-253-262-2009 -- (RCFCWCD) 10 images
- 2009 Oak Street Dam - RC-Oak-1-7-2009 -- (RCFCWCD) 7 images
- 2009 Oak Street Dam - RC-Oak-1-6-2009 -- (RCFCWCD) 6 images
- 2009 South Norco Channel - RC-South7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 South Norco Channel Lateral Line SB, Stage 3 - RC-South2-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Southwest Riverside Main Drain Pipe Line C - RC-SouthwestC-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Temescal Basin / Cajalco Lateral - RC-Temescal8-1-8-2009 -- (RCFCWCD) 2 images
- 2009 Temescal Channel - RC-Temescal7-1-8-2009 -- (RCFCWCD) 8 images
- 2009 Temescal Channel Coto Street - RC-Coto-1-8-2009 -- (RCFCWCD) 8 images
- 2009 Temescal Channel Coto Street - RC-Temescal-1-7-2009 -- (RCFCWCD) 7 images
- 2009 Temescal Detention Basin Cajalco Lateral - RC-Temescal2-1-8-2009 -- (RCFCWCD) 8 images
- 2009 West Norco Basin - RC-West-1-2-2009 -- (RCFCWCD) 2 images
- 2009 West Norco Basin - RC-West7-1-2-2009 -- (RCFCWCD) 2 images
- 2010 Mockingbird Canyon - RC-1-9-2010 -- (RCFCWCD) 9 images
- 2010 Mockingbird Canyon - RC-Mockingbird-1-8-2010 -- (RCFCWCD) 8 images
- 2010 Santa Ana River below Prado Dam - RC-1-33-2010 -- (RCFCWCD) 33 images
- 2010 Santa Ana River Phase II - RC-40-66-2010 -- (RCFCWCD) 27 images
- 2010 Santa Ana River Phase III - RC-67-79-2010 -- (RCFCWCD) 13 images
- 2010 Santa Ana River Phase III - RC-67-105-2010 -- (RCFCWCD) 39 images
- 2010 Santa Ana River Phase IV - RC-106-141-2010 -- (RCFCWCD) 36 images
- 2010 Temescal Creek - RC-Temescal-1-32-2010 -- (RCFCWCD) 32 images
- 2011 Brown Canyon Debris Basin - RC-Brown-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Brown Canyon Debris Basin - RC-Brown-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Day Creek Channel - RC-1-10-2011 -- (RCFCWCD) 10 images
- 2011 Eastvale Basin - RC-Eastvale-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Eastvale Basin - RC-Eastvale2-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Hunt Canyon Debris Basin - RC-Hunt-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Hunt Canyon Debris Basin - RC-Hunt-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Mabey Canyon Dam - RC-1-6-2011 -- (RCFCWCD) 6 images
- 2011 Main Street Dam - RC-Main-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Mockingbird Canyon Levee - RC-Mockingbird-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Norco Main Drain Pipeline Line S-1, Stage 1 - RC-1-28-2011 -- (RCFCWCD) 28 images
- 2011 Oak Street Dam Debris Basin - RC-Oak-1-3-2011 -- (RCFCWCD) 3 images
- 2011 South Norco Channel - RC-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Temescal Basin & Cajalco Lateral - RC-1-8-2011 -- (RCFCWCD) 8 images
- 2011 Temescal Channel Cota Street - RC-1-7-2011 -- (RCFCWCD) 7 images
- 2011 West Norco Basin - RC-West-1-2-2011 -- (RCFCWCD) 2 images
- 2012 Gavilan Hills Debris Basin - RC-Gavilan2-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Main Street Dam - RC-1-4-2012 -- (RCFCWCD) 4 images
- 2012 Mockingbird Canyon - RC-1-9-2012 -- (RCFCWCD) 9 images
- 2012 Norco Bluffs - RC-Norco-1-8-2012 -- (RCFCWCD) 8 images
- 2012 North Norco Channel, Line NB - RC-1-5-2012 -- (RCFCWCD) 5 images
- 2012 Oak Street Dam Debris Basin - RC-1-6-2012 -- (RCFCWCD) 6 images
- 2012 South Norco Channel Line SB Stage 3 - RC-1-3-2012 -- (RCFCWCD) 3 images
- 2012 South Norco Channel Stage 6 - RC-Norco6-1-4-2012 -- (RCFCWCD) 4 images
- 2012 Temescal Basin - RC-1-8-2012 -- (RCFCWCD) 8 images
- 2012 Temescal Channel - RC-1-7-2012 -- (RCFCWCD) 7 images
- 2012 West Norco Basin - RC-1-2-2012 -- (RCFCWCD) 2 images
- 2013 Brown Canyon Debris Basin - RC-Brown-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Eastvale Basin - RC-Eastvale-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Gavilan Hills -RC-Gavilan-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Hunt Canyon Debris Basin - RC-Hunt-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Mabey Canyon Dam - RC-Mabey-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Main Street Dam - RC-Main-1-4-2013 -- (RCFCWCD) 4 images
- 2013 Mockingbird Canyon - RC-Mockingbird-1-9-2013 -- (RCFCWCD) 9 images
- 2013 Norco Bluffs - RC-1-8-2013 -- (RCFCWCD) 8 images
- 2013 Norco Main Drain Pipeline Line NA - RC-NorcoNA-1-7-2013 -- (RCFCWCD) 7 images
- 2013 Oak Street Dam Debris Basin - RC-Oak-1-6-2013 -- (RCFCWCD) 6 images
- 2013 South Norco Channel - RC-South-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Temescal Basin & Cajalco Lateral - RC-Temescal2-1-8-2013 -- (RCFCWCD) 8 images
- 2013 Temescal Channel - RC-Temescal-1-7-2013 -- (RCFCWCD) 7 images
- 2013 West Norco Basin - RC-West-1-2-2013 -- (RCFCWCD) 2 images
- 2014 El Cerrito Channel Restoration - RC-1-6-2014 -- (RCFCWCD) 6 images
- 2015 Brown Canyon - RC-Brown-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Coldwater Canyon Wash - RC-Coldwater-1-13-2015 -- (RCFCWCD) 13 images
- 2015 Eastvale Basin - RC-Eastvale-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Gavilan Hills Debris Basin - RC-Gavilan-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Hunt Canyon Debris Basin - RC-Hunt-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Main Street Dam - RC-Main-1-5-2015 -- (RCFCWCD) 5 images
- 2015 Mockingbird Canyon- RC-1-9-2015 -- (RCFCWCD) 9 images
- 2015 Norco Bluffs- RC-Norco-1-8-2015 -- (RCFCWCD) 8 images
- 2015 Oak Street Dam - RC-Oak-1-6-2015 -- (RCFCWCD) 6 images
- 2015 South Norco Basin - RC-South2-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Temescal Basin & Cajalco Lateral - RC-1-8-2015 -- (RCFCWCD) 8 images
- 2015 Temescal Channel (Cota Street) - RC-Cota-1-7-2015 -- (RCFCWCD) 7 images
- 2015 West Norco Basin - RC-West-1-2-2015 -- (RCFCWCD) 2 images
-
Hemet (10,856)
The cities of Hemet, Idyllwild, and San Jacinto are located in the San Jacinto Valley of the southwestern region of Riverside County. Diamond Valley Lake is also in this area.
- 1938 San Jacinto River - Flight 5405-1938 -- (Rowe) 5 images
- 1939 Hemet Area - RC-AXM-18-FA-8-1939 -- (RCFCWCD) 11 images
- 1939 Idylwild - Flight 5750 - AXM-270-53-55 -- (Rowe) 12 images
- 1939 San Jacinto - AMX-207-26-111 -- (Rowe) 18 images
- 1939 San Jacinto - AXM-208-1-114-- (Rowe) 12 images
- 1948 Hemet -RC-30-93-1948 -- (RCFCWCD) 35 images
- 1948 Near Hemet -RC-1-33-1948 -- (RCFCWCD) 33 images
- 1948 San Jacinto River - RC-13-53-1948 -- (RCFCWCD) 12 images
- 1948 San Jacinto River - Bautista Creek - RC-1-53-1948 -- (RCFCWCD) 47 images
- 1949 Hemet Book 8A - RC-AXM-F-12F-30-170-1949 -- (RCFCWCD) 13 images
- 1949 Hemet Book 8A - RC-AXM-F-16F-31-41-1949 -- (RCFCWCD) 6 images
- 1949 Hemet Book 9A - RC-AXM-F-13F-42-190-1949 -- (RCFCWCD) 36 images
- 1949 Hemet Book 9A - RC-AXM-F-14F-55-213-1949 -- (RCFCWCD) 41 images
- 1949 Hemet Book 9A - RC-AXM-F-18F-80-210-1949 -- (RCFCWCD) 29 images
- 1949 Valle Vista Oblique -RC-22-31-1949 -- (RCFCWCD) 10 images
- 1957 Hemet - RC-57-69 -- (RCFCWCD) 13 images
- 1958 Hemet - RC-43-56 -- (RCFCWCD) 13 images
- 1958 Little Lake - RC-5-22-35 -- (RCFCWCD) 14 images
- 1958 San Jacinto & Bautista Creek - RC-1-1-15 -- (RCFCWCD) 15 images
- 1958 San Jacinto Percolation Test - RC-1-24 -- (RCFCWCD) 24 images
- 1959 San Jacinto & Valley -RC-36-59 -- (RCFCWCD) 24 images
- 1960 Bautista Creek - RC-1-87 -- (RCFCWCD) 87 images
- 1960 Hemet - RC-58-71-1960 -- (RCFCWCD) 14 images
- 1960 Idyllwild - RC-12-24 -- (RCFCWCD) 13 images
- 1960 San Jacinto River - RC-30-41 -- (RCFCWCD) 11 images
- 1961 Pico Road- RC-2-1-2 -- (RCFCWCD) 61 images
- 1961 San Jacinto - RC-38-49 -- (RCFCWCD) 11 images
- 1961 Winchester Area - RC-39-72 -- (RCFCWCD) 34 images
- 1962 Diamond Valley - RC-31-57 -- (RCFCWCD) 27 images
- 1962 San Jacinto - RC-21-80 -- (RCFCWCD) 27 images
- 1963 Hemet & State Street - RC-29-52-1963 -- (RCFCWCD) 24 images
- 1963 Hurkey Creek - RC-4-10-1963 -- (RCFCWCD) 7 images
- 1963 Lake Hemet - RC-3-7-1963 -- (RCFCWCD) 5 images
- 1963 San Jacinto-Soboba - RC-41-82-1963 -- (RCFCWCD) 42 images
- 1966 Bautista Canyon - RC-1-75-1966 -- (RCFCWCD) 74 images
- 1966 Idyllwild - Hurkey Creek - RC-3-5-1966 -- (RCFCWCD) 3 images
- 1966 San Jacinto River Flood Damage - RC-1-150-1966 -- (RCFCWCD) 98 images
- 1967 Bautista Canyon - RC-1-7-10-25 -- (RCFCWCD) 7 images
- 1967 Bautista Canyon - RC-17-85-1967 -- (RCFCWCD) 68 images
- 1967 Lakeview Flood Damage - RC-1-24-1967 -- (RCFCWCD) 24 images
- 1968 Idylwild - RC-1-2-1968 -- (RCFCWCD) 2 images
- 1968 Poppett Flat - RC-1-A-7-1968 -- (RCFCWCD) 9 images
- 1968 San Jacinto-Soboba - RC-1-14-1968 -- (RCFCWCD) 14 images
- 1968 San Jacinto - RC-1-20-1968 -- (RCFCWCD) 19 images
- 1969 Idylwild - RC-1-18-1969 -- (RCFCWCD) 14 images
- 1969 Lakeview - RC-134-139-1969 -- (RCFCWCD) 6 images
- 1969 Potrero Springs -RC-1-8-1969 -- (RCFCWCD) 8 images
- 1969 Salt Creek - RC-180-203-1969 -- (RCFCWCD) 22 images
- 1969 San Jacinto - RC-1-24-1969 -- (RCFCWCD) 24 images
- 1969 Stetson Ave - Hemet - RC-1-13-1969 -- (RCFCWCD) 13 images
- 1970 Hemet Landfill - RC-5-7-1970 -- (RCFCWCD) 3 images
- 1970 South Hemet (Cactus Valley) - RC-4-10-1970 -- (RCFCWCD) 7 images
- 1971 Idyllwild - RC-15-20-1971 -- (RCFCWCD) 6 images
- 1971 Palm Ave - RC-51-67-1971 -- (RCFCWCD) 17 images
- 1971 Reinhart Canyon - RC-1-1-5-3-1971 -- (RCFCWCD) 31 images
- 1971 Reinhart Canyon - RC-1-9-5-6-1971 -- (RCFCWCD) 36 images
- 1971 Saint Johns Canyon - RC-7-14-1971 -- (RCFCWCD) 8 images
- 1971 Whittier Avenue - RC-12-49-1971 -- (RCFCWCD) 38 images
- 1972 Domenigoni Valley - RC-55-104-1972 -- (RCFCWCD) 50 images
- 1972 Hemet Revision - RC-1-40-1972 -- (RCFCWCD) 38 images
- 1972 Valle Vista - RC-1-4-1972 -- (RCFCWCD) 4 images
- 1972 Winchester - RC-6-1972 -- (RCFCWCD) 1 image
- 1972 Winchester - RC-7-16-1972 -- (RCFCWCD) 10 images
- 1972 Winchester - RC-49-100-1972 -- (RCFCWCD) 2 images
- 1972 Winchester Low Obliques - RC-1-2-1972 -- (RCFCWCD) 2 images
- 1973 Bautista Creek - RC-1-3-1973 -- (RCFCWCD) 3 images
- 1973 Lakeview - RC-1-1-1-34-1973 -- (RCFCWCD) 34 images
- 1973 Mountain Avenue - RC-83-99-1973 -- (RCFCWCD) 17 images
- 1973 San Jacinto -RC-1-98-1973 -- (RCFCWCD) 98 images
- 1973 San Jacinto Drain - RC-7-17-1973 -- (RCFCWCD) 98 images
- 1973 St Johns Canyon - RC-100-105-1973 -- (RCFCWCD) 6 images
- 1974 Idyllwild Dumpsite - RC-3-5-1974 -- (RCFCWCD) 3 images
- 1974 Idyllwild Dumpsite Convergence - RC-Idyllwild-1-2-1974 -- (RCFCWCD) 2 images
- 1974 SJ Area - San Jacinto Reservoir - RC-1-31-1974 -- (RCFCWCD) 31 images
- 1974 SJ Area - San Jacinto Reservoir - RC-32-52-1974 -- (RCFCWCD) 21 images
- 1975 Antelope Valley -RC-12-30-1975 -- (RCFCWCD) 19 images
- 1975 Lamb Canyon - RC-3-6-1975 -- (RCFCWCD) 4 images
- 1976 Idylwild Disposal - RC-Idylwild-1-5-1976 -- (RCFCWCD) 5 images
- 1977 Double Butte Disposal - RC-Double-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Idylwild Disposal - RC-Idylwild-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Lamb Canyon Disposal - RC-1-5-1977 -- (RCFCWCD) 5 images
- 1977 San Jacinto River - RC-1-15-1977 -- (RCFCWCD) 15 images
- 1977 Sun City - RC-1-62-1977 -- (RCFCWCD) 62 images
- 1978 Idylwild Disposal - RC-Idyllwild-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Lamb Canyon Disposal - RC-Lamb-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Mead Valley Disposal - RC-Mead-1-3-1978 -- (RCFCWCD) 2 images
- 1978 San Jacinto River - RC-1-136-1978 -- (RCFCWCD) 136 images
- 1979 Double Butte Obliques - RC-3-5-9-1979 -- (RCFCWCD) 5 images
- 1979 Kitching Street Channel - RC-1-80-1979 -- (RCFCWCD) 80 images
- 1979 Kitching Street Channel - RC-Kitching-1-10-1979 -- (RCFCWCD) 10 images
- 1979 Lamb Canyon - RC-Lamb-1-6-1979 -- (RCFCWCD) 6 images
- 1979 Mead Valley Sanitary Landfill - RC-3-5-9-1979 -- (RCFCWCD) 10 images
- 1979 Nuevo Channel - RC-1-44-1979 -- (RCFCWCD) 39 images
- 1979 Paloma Valley - RC-1-39-1979 -- (RCFCWCD) 39 images
- 1979 Sunnymead - RC-1-44-1979 -- (RCFCWCD) 44 images
- 1980 Cactus Valley - RC-90-101-1980 -- (RCFCWCD) 12 images
- 1980 Diamond Valley - RC-102-121-1980 -- (RCFCWCD) 20 images
- 1980 Domenigoni - RC-122-139-1980 -- (RCFCWCD) 18 images
- 1980 Double Butte - RC-3-7-1980 -- (RCFCWCD) 3 images
- 1980 Edgemont Ortho - RC-1-34-1980 -- (RCFCWCD) 34 images
- 1980 Gavilan - RC-156-172-1980 -- (RCFCWCD) 17 images
- 1980 Hemet Channel - RC-1-35-1980 -- (RCFCWCD) 35 images
- 1980 Idyllwild - RC-Idyllwild-1-4-1980 -- (RCFCWCD) 4 images
- 1980 Menifee Valley - RC-140-155-1980 -- (RCFCWCD) 16 images
- 1980 Paloma Valley - RC-1-141-1980 -- (RCFCWCD) 141 images
- 1980 Reinhart Canyon - RC-36-41-1980 -- (RCFCWCD) 6 images
- 1980 Reinhart Canyon - RC-Reinhart-1-14-1980 13 imaages
- 1980 Salt Creek - RC-42-89-1980 -- (RCFCWCD) 48 images
- 1980 San Jacinto - RC-1-116-1980 -- (RCFCWCD) 116 images
- 1980 San Jacinto II - RC-117-125-1980 -- (RCFCWCD) 9 images
- 1980 San Jacinto River Flood Damage - RC-1-203-1980 -- (RCFCWCD) 203 images
- 1980 San Jacinto River Restoration Work - RC-1-54-1980 -- (RCFCWCD) 54 images
- 1980 West Hemet - RC-1-18-1980 -- (RCFCWCD) 15 images
- 1980 West Hemet Flood Damage - RC-19-145-1980 -- (RCFCWCD) 126 images
- 1981 Homeland/Lakeview Flooding - RC-1-82-1980 -- (RCFCWCD) 80 images
- 1982 Bautista Creek - RC-1-14-1982 -- (RCFCWCD) 14 images
- 1982 Green Acres - RC-1-15-1982 -- (RCFCWCD) 10 images
- 1982 Hemet Ortho - RC-1-70-1982 -- (RCFCWCD) 68 images
- 1982 Hemet Ortho - RC-10-105-1982 -- (RCFCWCD) 96 images
- 1982 Romoland - RC-1-61-1982 -- (RCFCWCD) 61 images
- 1982 San Jacinto Levee - RC-Jacinto-1-11-1982 -- (RCFCWCD) 11 images
- 1982 San Jacinto River - RC-1-22-1982 -- (RCFCWCD) 22 images
- 1983 Double Butte Sanitary Landfill - RC-Double-1-9-1983 -- (RCFCWCD) 9 images
- 1983 Double Butte Sanitary Landfill - RC-Double-1-10-1983 -- (RCFCWCD) 10 images
- 1983 Hemet - Elsinore - RC-Hemet-1-16-1983 -- (RCFCWCD) 16 images
- 1983 Idylwild Sanitary Landfill - RC-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Idylwild - RC-Idylwild-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Lamb Canyon Sanitary Landfill - RC-Lamb-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Lamb Canyon Sanitary Landfill - RC-Lamb-1-6-1983 -- (RCFCWCD) 6 images
- 1983 San Jacinto River - RC-40-103-1983 -- (RCFCWCD) 63 images
- 1983 San Jacinto River - RC-104-123-1983 -- (RCFCWCD) 20 images
- 1983 Winchester Ortho - RC-1-57-1983 -- (RCFCWCD) 57 images
- 1983 Winchester Ortho - RC-63-78-1983 -- (RCFCWCD) 16 images
- 1984 Alessandro Basin -RC-Alessandro-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Bautista Basin - RC-Bautista-1-3-1984 -- (RCFCWCD) 3 images
- 1984 Double ButteSanitary Landfill - RC-Double-1-13-1984 -- (RCFCWCD) 13 images
- 1984 Idylwild -RC-Idylwild1-1-2-1984 -- (RCFCWCD) 2 images
- 1984 Idylwild Sanitary Landfill - RC-Idylwild2-1-2-1984 -- (RCFCWCD) 2 images
- 1984 Idylwild Sanitary Landfill - RC-1-2-1984 -- (RCFCWCD) 2 images
- 1984 San Jacinto River - RC-1-51-1984 -- (RCFCWCD) 49 images
- 1984 Keen Camp Sanitary Landfill - RC-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Salt Creek Channel - RC-1-85-1984 -- (RCFCWCD) 85 images
- 1984 Salt River Channel Phase II -RC-1-47-1984 -- (RCFCWCD) 47 images
- 1984 San Jacinto Drain - Buena Vista Basin - RC-Jacinto-1-15-1984 -- (RCFCWCD) 15 images
- 1984 San Jacinto River Park - RC-3-7-1984 -- (RCFCWCD) 5 images
- 1984 Sunnymead Channel - RC-1-17-1984 -- (RCFCWCD) 17 images
- 1984 Sunnymead Channel Part 1 - RC-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Sunnymead Storm Drain - RC-1-22-1984 -- (RCFCWCD) 22 images
- 1985 Hemet A-3 to Acacia - RC-1-10-1985 -- (RCFCWCD) 10 images
- 1985 Salt Creek - RC-Salt-1-17-1985 -- (RCFCWCD) 17 images
- 1985 San Sevaine Channel - RC-1-26-1985 -- (RCFCWCD) 26 images
- 1985 Sunnymead Line Q - RC-1-15-1985 -- (RCFCWCD) 15 images
- 1986 Mead Valley Outlet Channel -RC-Mead-1-7-1986 -- (RCFCWCD) 7 images
- 1986 San Jacinto MPD Line E-3 - RC-Jacinto-1-4-1986 -- (RCFCWCD) 4 images
- 1986 Stetson Avenue Channel - RC-Stetson-1-13-1986 -- (RCFCWCD) 13 images
- 1987 Bautista Creek Channel - RC-1-34-1987 -- (RCFCWCD) 32 images
- 1987 Little Lake Basin - RC-Lake-1-2-1987 -- (RCFCWCD) 2 images
- 1987 Little San Gorgonio - RC-SG-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Metz Basin - RC-Metz-1-5-1987 -- (RCFCWCD) 5 images
- 1987 Parkhill Basin - RC-Parkhill-1-2-1987 -- (RCFCWCD) 2 images
- 1987 Perris & Third Street Basin - RC-Perris-1-3-1987 -- (RCFCWCD) 3 images
- 1987 Valle Vista Ortho - RC-1-32-1987 -- (RCFCWCD) 32 images
- 1987 Valle Vista Ortho - RC-1-20-1987 -- (RCFCWCD) 20 images
- 1988 Bautista - RC-Bautista-1-5-1988 -- (RCFCWCD) 5 images
- 1988 Double Butte Spot - RC-1-13-1988 -- (RCFCWCD) 13 images
- 1988 Lamb Canyon - RC-1-10-1988 -- (RCFCWCD) 10 images
- 1988 Lamb Canyon - RC-Lamb-1-1988 -- (RCFCWCD) 1 images
- 1988 Little Lake Basin - RC-Little-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Mead Calley - RC-1-9-1988 -- (RCFCWCD) 9 images
- 1988 Mead Valley Spot - RC-Mead-1-1988 -- (RCFCWCD) 1 image
- 1988 Metz Basin - RC-Metz-1-5-1988 -- (RCFCWCD) 5 images
- 1988 Parkhill Basin - RC-Parkhill-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Perris Third Street Basin - RC-1-3-1988 -- (RCFCWCD) 3 images
- 1988 Pigeon Pass Basin - RC-Pigeon-1-6-1988 -- (RCFCWCD) 6 images
- 1988 San Jacinto River - RC-50-127-1988 -- (RCFCWCD) 78 images
- 1988 Stetson Avenue Channel - RC-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Sunnymead Line B-8 and B-11 - RC-Sunnymead-1-21-1988 -- (RCFCWCD) 21 images
- 1988 Sunnymead MDP Line B-5 - RC-Sunnymead-1-2-1988 -- (RCFCWCD) 2 images
- 1989 Buena Vista - RC-Buena-1-3-1989 -- (RCFCWCD) 3 images
- 1989 Idylwild or Keen Camp - RC-Idylwild-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Little Lake Basin - RC-Little-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Little Lake Line-B - RC-1-60-1989 -- (RCFCWCD) 60 images
- 1989 Mead Valley - RC-1-7-1989 -- (RCFCWCD) 7 images
- 1989 Mead Valley Spot - RC-Mead-1-1989 -- (RCFCWCD) 1 image
- 1989 Metz Basin - RC-Metz-1-5-1989 -- (RCFCWCD) 5 images
- 1989 Parkhill Basin - RC-Parkhill-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Perris Third Street Basin - RC-Perris-1-3-1989 -- (RCFCWCD) 3 images
- 1989 San Jacinto Extension - RC-Jacinto-1-6-1989 -- (RCFCWCD) 6 images
- 1989 Sunnymead Line H-7 - RC-Sunnymead-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Winchester -D - RC-Winchester-1-4-1989 -- (RCFCWCD) 4 images
- 1990 Baristo Wash - RC-Baristo-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Bradley Road Channel - RC-1-15-1990 -- (RCFCWCD) 15 images
- 1990 Canyon Springs Basin - RC-Canyon-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Gilman Springs Road - RC-1-29-1990 -- (RCFCWCD) 29 images
- 1990 Hemet - RC-Hemet-1-5-1990 -- (RCFCWCD) 5 images
- 1990 Lamb Canyon - RC-1-17-1990 -- (RCFCWCD) 17 images
- 1990 Lasselle Basin - RC-Lasselle-1-2-90 -- (RCFCWCD) 4 images
- 1989 Winchester -D - RC-Winchester-1-4-1989 -- (RCFCWCD) 4 images
- 1990 Bautista Creek - RC-Bautista-1-5-1990 -- (RCFCWCD) 6 images
- 1990 Metz Basin - RC-Metz-1-5-1990 -- (RCFCWCD) 5 images
- 1990 Parkhill Basin - RC-Parkhill-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Perris Third Street Basin - RC-Perris-1-3-1990 -- (RCFCWCD) 2 images
- 1990 Little Basin - RC-Little-1-2-1990 -- (RCFCWCD) 2 images
- 1990 San Jacinto River - RC-1-72-1990 -- (RCFCWCD) 72 images
- 1991 Buena Vista - RC-Buena-1-3-1991 -- (RCFCWCD) 3 images
- 1991 Park Hill Basin - RC-Park-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Perris Third Street - RC-Perris-1-3-1991-- (RCFCWCD) 3 images
- 1991 San Jacinto Drain - RC-Jacinto-1-9-1991 -- (RCFCWCD) 9 images
- 1991 San Jacinto River - RC-1-106-1991 -- (RCFCWCD) 106 images
- 1991 San Jacinto River - RC-1-40-1991 -- (RCFCWCD) 40 images
- 1991 San Jacinto River - RC-1-114-1991 -- (RCFCWCD) 114 images
- 1992 Double Butte - RC-1,2,4,6,25-27-1992 -- (RCFCWCD) 7 images
- 1992 Lamb Canyon Sanitary Landfill - RC-1-32-1992 -- (RCFCWCD) 32 images
- 1992 Lamb Canyon Sanitary Landfill Spot - RC-Lamb-A-1992 -- (RCFCWCD) 1 image
- 1992 Lamb 2 - Lamb2-A-1992 -- (RCFCWCD) 1 image
- 1992 Metz Basin - RC-Metz-1-5-1992 -- (RCFCWCD) 5 images
- 1992 Perris Valley Lateral B - RC-1-13-1992 -- (RCFCWCD) 13 images
- 1992 Sunnymead P-2 - RC-1-27-1992 -- (RCFCWCD) 27 images
- 1993 Bautista Basin - RC-Bautista-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Bautista Creek - RC-1-15-1993 -- (RCFCWCD) 15 images
- 1993 Hemet - RC-1-364-1993-1 -- (RCFCWCD) 252 images
- 1993 Hemet - RC-1-364-1993-2 -- (RCFCWCD) 112 images
- 1993 Homeland - RC-1-36-1993 -- (RCFCWCD) 36 images
- 1993 Little Lake Pit - RC-Little-1-2-1993 -- (RCFCWCD) 2 images
- 1993 San Jacinto - RC-1-146-1993 -- (RCFCWCD) 146 images
- 1993 San Jacinto - RC-1-26-1993 -- (RCFCWCD) 26 images
- 1993 San Jacinto-Lakeview - RC-1-7-1993 -- (RCFCWCD) 7 images
- 1993 San Jacinto Line C and Lateral 5 - RC-Jacinto-1-4-1993 -- (RCFCWCD) 4 images
- 1994 Bautista Basin - RC-1-11-1994 -- (RCFCWCD) 12 images
- 1994 Bautista Basin - RC-Bautista-3-1994 -- (RCFCWCD) 1 image
- 1994 Double Butte Sanitary Landfill - RC-1-16-1994 -- (RCFCWCD) 16 images
- 1994 Double Butte Sanitary Landfill - RC-Double-1-16-1994 -- (RCFCWCD) 16 images
- 1994 Double Butte Spot - RC-Double-A-1994 -- (RCFCW 8 images
- 1994 Hemet Sanitary Landfill - RC-Hemet2-1-3-1994 -- (RCFCWCD) 3 images
- 1994 Hemet Spot - RC-Hemet-A-1994 -- (RCFCWCD) 1 image
- 1994 Idyllwild Sanitary Landfill - RC-Idyllwild2-1-3-1994 -- (RCFCWCD) 3 images
- 1994 Idyllwild Spot - RC-Idyllwild-A-1994 -- (RCFCWCD) 1 image
- 1994 Lamb Canyon LF Equipment - RC-Lamb-1-2-1994 -- (RCFCWCD) 2 images
- 1994 Lamb Canyon Sanitary Landfill - RC-Lamb3-1-16-1994 -- (RCFCWCD) 17 images
- 1994 Lamb Spot - RC-Lamb2-A-1994 -- (RCFCWCD) 1 image
- 1994 Mead Valley Outlet Channel - RC-Mead-1-15-1994 -- (RCFCWCD) 15 images
- 1994 Mead Valley Sanitary Landfill - RC-1-10-1994 -- (RCFCWCD) 10 images
- 1994 Mead Valley Spot - RC-Mead-A-1994 -- (RCFCWCD) 1 image
- 1994 Perris Valley - RC-1-25-1994 -- (RCFCWCD) 25 images
- 1994 San Jacinto River - RC-1-72-1994 -- (RCFCWCD) 72 images
- 1994 San Jacinto River - RC-1-256-1994 -- (RCFCWCD) 252 images
- 1994 Sunnymead H7 and H9 - RC-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Valley Vista Ortho - RC-25-36-1994 -- (RCFCWCD) 12 images
- 1994 Valley Vista Ortho - RC-1-24-1994 -- (RCFCWCD) 36 images
- 1994 Valley Vista Ortho - RC-1-41-1994 -- (RCFCWCD) 41 images
- 1995 Bautista Court Exhibit - RC-Bautista-1-6-1995 -- (RCFCWCD) 6 images
- 1995 Bautista Inlet - RC-Bautista-1-13-1995 -- (RCFCWCD) 13 images
- 1995 Bradley Road Channel - RC-Bradley-1-2-1995 -- (RCFCWCD) 2 images
- 1995 Buena Vista Basin - RC-Buena-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Double Butte Sanitary Landfill - RC-Double-1-14-1995 -- (RCFCWCD) 14 images
- 1995 Hemet Sanitary Landfill - RC-Hemet2-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Hemet Sanitary Landfill Spot - RC-Hemet-A-1995 -- (RCFCWCD) 1 image
- 1995 Idylwild Sanitary Landfill - RC-Idylwild2-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Idylwild Sanitary Landfill Spot - RC-Idylwild-A-1995 -- (RCFCWCD) 1 image
- 1995 Lamb Canyon Sanitary Landfill - RC-1-16-1995 -- (RCFCWCD) 16 images
- 1995 Lamb Sanitary Landfill Spot - RC-Lamb-A-1995 -- (RCFCWCD) 1 image
- 1995 Mead Valley Sanitary Landfill - RC-1-10-1995 -- (RCFCWCD) 10 images
- 1995 Mead Valley Sanitary Landfill Spot - RC-Mead-A-1995 -- (RCFCWCD) 1 image
- 1995 Potrero Creek - RC-Potrero-1-14-1995 -- (RCFCWCD) 14 images
- 1995 San Jacinto Drain - RC-Jacinto-1-12-1995 -- (RCFCWCD) 12 images
- 1995 San Jacinto River - RC-1-60-1995 -- (RCFCWCD) 60 images
- 1995 Sunnymead Line B-7 - RC-Sunnymead-1-4-1995 -- (RCFCWCD) 4 images
- 1996 Double Butte - RC-Double-1-2-1996 -- (RCFCWCD) 2 images
- 1996 Double Butte Sanitary Landfill - RC-1-17-1996 -- (RCFCWCD) 16 images
- 1996 Hemet Sanitary Landfill - RC-Hemet-1-3-1996 -- (RCFCWCD) 3 images
- 1996 Hemet Ortho - RC-1-74-1996 -- (RCFCWCD) 74 images
- 1996 Idyllwild Sanitary Landfill - RC-Idyllwild-1-3-1996 -- (RCFCWCD) 3 images
- 1996 Lamb Canyon Sanitary Landfill Spot - RC-Lamb-A-1996 -- (RCFCWCD) 1 image
- 1996 Lamb Sanitary Landfill - RC-Lamb2-1-16-1996 -- (RCFCWCD) 16 images
- 1996 Mead Valley Sanitary Landfill - RC-1-9-1996 -- (RCFCWCD) 9 images
- 1996 Mead Valley Sanitary Landfill - RC-1-10-1996 -- (RCFCWCD) 10 images
- 1996 Mead Valley Sanitary Landfill Spot - RC-Mead-A-1996 -- (RCFCWCD) 1 image
- 1996 Mead Valley Sanitary Landfill Spo t- RC-Meade2-A-1996 -- (RCFCWCD) 1 image
- 1997 Double Butte Sanitary Landfill - RC-1-19-1997 -- (RCFCWCD) 19 images
- 1997 Double Butte Sanitary Landfill Spot - RC-Double-A-1997 -- (RCFCWCD) 1 image
- 1997 Hemet Channel - RC-1-47-1997 -- (RCFCWCD) 48 images
- 1997 Lakeview Dam - RC-Lakeview-1-11-1997 -- (RCFCWCD) 11 images
- 1997 Lamb Canyon Sanitary Landfill - RC-1-1-3-5-1997 -- (RCFCWCD) 12 images
- 1997 Lamb Canyon Sanitary Landfill Spot -RC-Lamb-1-1997 -- (RCFCWCD) 1 image
- 1997 Mead Valley Sanitary Landfill Spot - RC-Mead-1-1997 -- (RCFCWCD) 1 image
- 1997 Mead Valley Sanitary Landfill - Mead2-1-1-2-6-1997 -- (RCFCWCD) 13 images
- 1997 Nuevo Channel - RC-Nuevo-1-14-1997 -- (RCFCWCD) 14 images
- 1997 Portrero Creek - RC-Portrero-1-7-1997 -- (RCFCWCD) 7 images
- 1997 Potrero Basin - RC-Potrero-1-8-1997 -- (RCFCWCD) 8 images
- 1997 San Jacinto Drain - RC-Jacinto-1-11-1997 -- (RCFCWCD) 11 images
- 1997 San Jacinto Reservoir - RC-Jacinto2-1-12-1997 -- (RCFCWCD) 12 images
- 1997 San Jacinto River - RC-1-102-1997 -- (RCFCWCD) 101 images
- 1997 San Jacinto River - RC-1-102-1997 -- (RCFCWCD) 103 images
- 1997 San Jacinto River Stage 4 - RC-1-49-1997 -- (RCFCWCD) 49 images
- 1998 Bautista Basin - RC-Bautista-1-13-1998 -- (RCFCWCD) 13 images
- 1998 Hemet Channel - RC-Hemet-1-47-1998 -- (RCFCWCD) 47 images
- 1998 Nuevo Channel - RC-Nuevo-1-14-1998 -- (RCFCWCD) 14 images
- 1998 Perris Valley Channel; - RC-1-16-1998 -- (RCFCWCD) 16 images
- 1998 Potrero Creek Basin - RC-1-7-1998 -- (RCFCWCD) 7 images
- 1998 Potrero Creek Basin - RC-Potrero-1-14-1998 -- (RCFCWCD) 14 images
- 1998 San Jacinto Drain - RC-1-11-1998 -- (RCFCWCD) 11 images
- 1998 San Jacinto River - RC-1-104-1998 -- (RCFCWCD) 104 images
- 1998 San Jacinto Riverside Stage 4 - RC-1-14-1998 -- (RCFCWCD) 14 images
- 1999 Double Butte Sanitary Landfill - RC-1-17-1999 -- (RCFCWCD) 17 images
- 1999 Double Butte Sanitary Landfill Spot - RC-Double-A-1999 -- (RCFCWCD) 1 image
- 1999 Hemet Channel - RC-1-45-1999 -- (RCFCWCD) 45 images
- 1999 Lakeview Dam - RC-Lakeview-1-10-1999 -- (RCFCWCD) 10 images
- 1999 Lamb Canyon Sanitary Landfill - RC-1-21-1999 -- (RCFCWCD) 21 images
- 1999 Lamb Canyon Sanitary Landfill Spot - RC-Lamb-A-1999 -- (RCFCWCD) 1 image
- 1999 Mead Valley Sanitary Landfill - RC-1-12-1999 -- (RCFCWCD) 12 images
- 1999 Mead Valley Sanitary Landfill Spot - RC-Mead-A-1999 -- (RCFCWCD) 1 image
- 1999 Moreno Line F-1 - RC-Moreno-1-4-1999 -- (RCFCWCD) 4 images
- 1999 Nuevo Channel - RC-Nuevo-1-14-1999 -- (RCFCWCD) 14 images
- 1999 Perris Valley Channel - RC-Perris-1-16-1999 -- (RCFCWCD) 16 images
- 1999 Potrero Creek - RC-Potrero-1-7-1999 -- (RCFCWCD) 6 images
- 1999 San Jacinto Drain - RC-Jacinto-1-11-1999 -- (RCFCWCD) 11 images
- 1999 San Jacinto River - RC-1-106-1999 -- (RCFCWCD) 105 images
- 1999 Stetson Avenue Channel Stages 3 and 4 - RC-Stetson-1-16-1999 -- (RCFCWCD) 15 images
- 1999 Sunnymead Main Drain Pipeline - RC-Sunnymead-1-2-1999 -- (RCFCWCD) 2 images
- 2000 Bautista Creek Dam - RC-1-13-2000 -- (RCFCWCD) 13 images
- 2000 Hemet Channel - RC-1-49-2000 -- (RCFCWCD) 49 images
- 2000 Hemet Sanitary Landfill - RC-Hemet2-1-3-2000 -- (RCFCWCD) 3 images
- 2000 Hemet Sanitary Landfill Spot - RC-Hemet-A-2000 -- (RCFCWCD) 1 image
- 2000 Idyllwild Sanitary Landfill - RC-Idyllwild2-1-3-2000 -- (RCFCWCD) 3 images
- 2000 Idyllwild Sanitary L:andfill Spot - RC-Idyllwild-A-2000 -- (RCFCWCD) 1 image
- 2000 Lakeview Dam - RC-Lakeview-1-10-2000 -- (RCFCWCD) 10 images
- 2000 Lamb Canyon Sanitary Landfill - RC-Lamb-1-19-2000 -- (RCFCWCD) 19 images
- 2000 Lamb Canyon Sanitary Landfill - RC-1-19-2000 -- (RCFCWCD) 19 images
- 2000 Lamb Canyon Sanitary Landfill Spot - RC-Lamb-A-2000 -- (RCFCWCD) 1 image
- 2000 Mead Sanitary Landfill - RC-1-12-2000 -- (RCFCWCD) 12 images
- 2000 Mead Sanitary Landfill Spot - RC-Mead-A-2000 -- (RCFCWCD) 1 image
- 2000 Nuevo Channel - RC-Nuevo-1-14-2000 -- (RCFCWCD) 14 images
- 2000 Potrero Creek Basin - RC-1-7-2000 -- (RCFCWCD) 7 images
- 2000 Salt Creek Channel - RC-1-30-2000 -- (RCFCWCD) 30 images
- 2000 San Jacinto Drains - RC-1-11-2000 -- (RCFCWCD) 11 images
- 2000 San Jacinto River - RC-1-106-2000 -- (RCFCWCD) 106 images
- 2001 Double Butte Sanitary Landfill - RC-Double-1-17-2001 -- (RCFCWCD) 17 images
- 2001 Hemet Channel - RC-Hemet-1-47-2001 -- (RCFCWCD) 47 images
- 2001 Lakeview Dam - RC-1-10-2001 -- (RCFCWCD) 10 images
- 2001 Lamb Canyon - RC-Lamb-1-19-2001 -- (RCFCWCD) 19 images
- 2001 Lamb Canyon Sanitary Landfill - RC-1-19-2001 -- (RCFCWCD) 19 images
- 2001 Nuevo Channel - RC-Nuevo-1-15-2001 -- (RCFCWCD) 15 images
- 2001 Ortego Channel - RC-Ortego-1-4-2001 -- (RCFCWCD) 4 images
- 2001 Perris Valley Channel - RC-1-17-2001 -- (RCFCWCD) 17 images
- 2001 Potrero Creek Basin - RC-1-7-2001 -- (RCFCWCD) 7 images
- 2001 San Jacinto Drain - RC-Jacinto-1-11-2001 -- (RCFCWCD) 11 images
- 2001 San Jacinto River - RC-1-99-2001 -- (RCFCWCD) 99 images
- 2002 Bautista Basin - RC-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Double Butte Sanitary Landfill - RC-1-17-2002 -- (RCFCWCD) 17 images
- 2002 Lamb Canyon Sanitary Landfill - RC-1-20-2002 -- (RCFCWCD) 20 images
- 2002 Nuevo Channel - RC-Nuevo-1-6-2002 -- (RCFCWCD) 6 images
- 2002 Paloma Valley Ortho - RC-1-32-2002 -- (RCFCWCD) 32 images
- 2002 San Jacinto River - RC-1-72-2002 -- (RCFCWCD) 72 images
- 2003 Bachelor Mountain - RC-Bachelor-1-27-2003 -- (RCFCWCD) 27 images
- 2003 Hemet Channel - RC-1-36-2003 -- (RCFCWCD) 36 images
- 2003 Hemet Channel - RC-Hemet-1-24-2003 -- (RCFCWCD) 24 images
- 2003 Hemet Vernal Pools - RC-Hemet-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Hemet Vernal Pools - RC-Hemet2-7-2003 -- (RCFCWCD) 1 image
- 2003 Hemet Vernal Pools - RC-Hemet3-A-2003 -- (RCFCWCD) 1 image
- 2003 Hemet Vernal Pools - RC-Hemet4-1-6-2003 -- (RCFCWCD) 5 images
- 2003 Hemet Vernal Pools - RC-Hemet5-A-2003 -- (RCFCWCD) 1 image
- 2003 Hemet Vernal Pools - RC-Hemet6-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Lakeview Dam - RC-Lakeview-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Lakeview Dam - RC-Lakeview2-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Lamb Canyon Sanitary Landfill - RC-Lamb-1-20-2003 -- (RCFCWCD) 20 images
- 2003 Nuevo Channel - RC-Nuevo-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Potrero Creek - RC-Potrero-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Potrero Creek - RC-Potrero2-1-6-2003 -- (RCFCWCD) 6 images
- 2003 San Jacinto Line E - RC-Jacinto-RC-1-27-2003 -- (RCFCWCD) 27 images
- 2003 San Jacinto River - RC-1-34-2003 -- (RCFCWCD) 34 images
- 2003 San Jacinto River - RC-Jacinto2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 San Jacinto River North - RC-North-1-2003 -- (RCFCWCD) 1 image
- 2003 San Jacinto River Stage I - RC-San-1-3-2003 -- (RCFCWCD) 3 images
- 2003 San Jacinto River Stage I - RC-SJR3-1-2003 -- (RCFCWCD) 1 image
- 2003 San Jacinto River Stage I - RC-SJR-1-2003 -- (RCFCWCD) 1 image
- 2003 San Jacinto River Stage III - RC-San2-1-3-2003 -- (RCFCWCD) 3 images
- 2003 San Jacinto River Stage III - RC-SJR2-1-2-2003 -- (RCFCWCD) 2 image
- 2003 Ski Land - RC-Ski-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Ski Land - RC-Ski2-1-2003 -- (RCFCWCD) 1 image
- 2003 Ski Land - RC-Ski3-1-2003 -- (RCFCWCD) 1 image
- 2004 Hemet Channel - RC-Hemet-1-24-2004 -- (RCFCWCD) 22 images
- 2004 Hemet Main Drain Pipe Line - RC-1-1-5-3-2004 -- (RCFCWCD) 19 images
- 2004 Lakeview Dam - RC-Lakeview-1-4-2004 -- (RCFCWCD) 4 images
- 2004 Nuevo Channel - RC-Nuevo-1-6-2004 -- (RCFCWCD) 6 images
- 2004 Potrero Creek - RC-Potrero-1-6-2004 -- (RCFCWCD) 6 images
- 2004 Potrero Ortho - Rc-1-98-2004 -- (RCFCWCD) 98 images
- 2004 San Jacinto River - RC-1-34-2004 -- (RCFCWCD) 34 images
- 2004 San Jacinto River-Cedar - RC-1-71-2004 -- (RCFCWCD) 71 images
- 2004 San Jacinto River Cedar to Bridge Street - RC-1-71-2004 -- (RCFCWCD) 71 images
- 2004 San Jacinto River I-15 to Canyon Lake - RC-1-37-2004 -- (RCFCWCD) 37 images
- 2004 San Jacinto River Cedar to Bridge Street - RC-1-71-2004 -- (RCFCWCD) 71 images
- 2005 Bautista Basin - RC-Bautista-1-5-2005 -- (RCFCWCD) 3 images
- 2005 Cactus/Heacock - RC-1-15-2005 -- (RCFCWCD) 16 images
- 2005 French Valley - RC-1-11-2005 -- (RCFCWCD) 11 images
- 2005 Heacock & Cactus Channels - RC-1-28-2005 -- (RCFCWCD) 28 images
- 2005 Hemet Channel - RC-Hemet7-1-24-2005 -- (RCFCWCD) 24 images
- 2005 Lakeview - RC-1-99-2005 -- (RCFCWCD) 100 images
- 2005 Nuevo Channel - RC-Nuevo7-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Potrero Basin - RC-Potrero-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Portrero Creek Basin - RC-Portrero7-1-6-2005 -- (RCFCWCD) 6 images
- 2005 San Jacinto Stage 3 - RC-1-32-2005 -- (RCFCWCD) 32 images
- 2005 San Jacinto Drain - RC-Jacinto7-1-9-2005 -- (RCFCWCD) 9 images
- 2005 San Jacinto Drain - RC-Jacinto8-1-9-2005 -- (RCFCWCD) 9 images
- 2005 San Jacinto River Cedar to Bridge Street - RC-Jacinto9-1-71-2005 -- (RCFCWCD) 71 images
- 2005 West Hemet/East Winchester - RC-1-50-2005 -- (RCFCWCD) 50 images
- 2006 Agriscape - RC-Agriscape-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Hemet Channel - RC-1-24-2006 -- (RCFCWCD) 24 images
- 2006 Hemet Main Drain Pipe Line D Stage 5 - RC-Hemet-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Palomar Detention Basin - RC-Palomar-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Potrero Basin - RC-Potrero-1-6-2006 -- (RCFCWCD) 6 images
- 2006 San Jacinto Drain - RC-Jacinto-1-10-2006 -- (RCFCWCD) 10 images
- 2006 San Jacinto River - RC-San-SJ-1-36-2006 -- (RCFCWCD) 36 images
- 2006 San Jacinto River - RC-1-34-2006 -- (RCFCWCD) 34 images
- 2007 Lamb Canyon - RC-1-25-2007 -- (RCFCWCD) 25 images
- 2007 Little Lake Basin - RC-Little-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Nuevo Basin - RC-Nuevo-1-6-2007 -- (RCFCWCD) 6 images
- 2007 San Jacinto Drain - RC-Jacinto2-1-9-2007 -- (RCFCWCD) 4 images
- 2007 San Jacinto River COE-BRG - RC-1-71-2007 -- (RCFCWCD) 71 images
- 2007 San Jacinto Stage 4 - RC-1-57-2007 -- (RCFCWCD) 57 images
- 2008 Little Lake Basin - RC-Little7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Mead Valley Sanitary Landfill - RC-Mead7-1-12-2008 -- (RCFCWCD) 12 image
- 2008 Nuevo Channel - RC-Nuevo7-1-6-2008 -- (RCFCWCD) 6 images
- 2008 Potrero Creek Basin - RC-Potrero2-1-6-2008 -- (RCFCWCD) 6 images
- 2008 Potrero Basin - RC-1-6-2008 -- (RCFCWCD) 6 images
- 2008 San Jacinto Line E - RC-1-13-2008 -- (RCFCWCD) 12 images
- 2008 San Jacinto River - RC-1-70-2008 -- (RCFCWCD) 70 images
- 2008 San Jacinto River San Jacinto to Bridge - RC-1-31-2008 -- (RCFCWCD) 31 images
- 2008 San Jacinto River I-215 to Canyon Lake - RC-1-34-2008 -- (RCFCWCD) 34 images
- 2008 San Jacinto River Levee - RC-1-66-2008 -- (RCFCWCD) 66 images
- 2009 Bautista Basin - RC-Bautista7-1-3-2009 -- (RCFCWCD) 3 image
- 2009 Hemet Channel - RC-Hemet-1-24-2009 -- (RCFCWCD) 24 images
- 2009 Hemet Channel - RC-Hemet7-1-24-2009 -- (RCFCWCD) 24 images
- 2009 Little Lake Basin - RC-Little8-1-2-2009 -- (RCFCWCD) 2 image
- 2009 Nuevo Channel - RC-Nuevo7-1-1-2009 -- (RCFCWCD) 6 image
- 2009 Palomar Detention Basin - RC-Palomar-1-2-2009 -- (RCFCWCD) 2 images
- 2009 San Jacinto River - RC-1-70-2009 -- (RCFCWCD) 70 image
- 2009 San Jacinto Line E - RC-SanE-1-13-2009 -- (RCFCWCD) 13 images
- 2009 San Jacinto Line E - RC-1-13-2009 -- (RCFCWCD) 13 images
- 2009 San Jacinto Main Drain Pipe Line C4 & C5 - RC-Jacinto7-1-11-2009 -- (RCFCWCD) 11 images
- 2009 San Jacinto River Stage 4 - RC-1-10-2009 -- (RCFCWCD) 10 images
- 2010 Hemet Main Drain Pipe Line C / Stage 4 - RC-1-13-2010 -- (RCFCWCD) 13 images
- 2010 Lamb Canyon Sanitary Landfill - RC-Lamb-1-22-2010 -- (RCFCWCD) 22 images
- 2010 Lamb Canyon Spot - RC-Spot-1-2010 -- (RCFCWCD) 1 image
- 2010 Romoland - RC-1-19-2010 -- (RCFCWCD) 19 images
- 2010 San Jacinto River - RC-1-23-2010 -- (RCFCWCD) 23 images
- 2010 San Jacinto River I-215 to Canyon Lake - RC-1-34-2010 -- (RCFCWCD) 34 images
- 2010 San Jacinto to French Valley - RC-1-132-2010 -- (RCFCWCD) 132 images
- 2010 San Jacinto to French Valley Obliques - RC-1-3-2010 -- (RCFCWCD) 3 images
- 2011 Bautista Basin - RC-Bautista-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Double Butte Sanitary Landfill - RC-1-17-2011 -- (RCFCWCD) 17 images
- 2011 Double Butte Sanitary Landfill - RC-Double-A-2011 -- (RCFCWCD) 1 image
- 2011 Hemet Channel - RC-1-24-2011 -- (RCFCWCD) 24 images
- 2011 Little Lake Basin - RC-Little2-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Little Lake Main Drain Pipeline - RC-1-15-2011 -- (RCFCWCD) 15 images
- 2011 Nuevo Channel - RC-Nuevo-1-6-2011 -- (RCFCWCD) 6 images
- 2011 San Jacinto - RC-24-68-2011 -- (RCFCWCD) 45 images
- 2011 San Jacinto River Army Corp of Engineers to Bridge - RC-1-70-2011 -- (RCFCWCD) 70 images
- 2011 San Jacinto Line E - RC-JacintoE-1-13-2011 -- (RCFCWCD) 13 images
- 2012 Bautista Basin - RC-Bautista7-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Hemet Channel - RC-1-24-2012 -- (RCFCWCD) 24 images
- 2012 Hemet Main Drain Pipeline Line D, Stage 7 - RC-Hemet7-1-5-2012 -- (RCFCWCD) 5 images
- 2012 Nuevo Channel - RC-Nuevo7-1-6-2012 -- (RCFCWCD) 6 images
- 2012 San Jacinto Line E - RC-SanE-1-13-2012-- (RCFCWCD) 13 images
- 2012 San Jacinto River to Army Corp of Engineers to Bridge - RC-1-70-2012 -- (RCFCWCD) 70 images
- 2012 San Jacinto Riverside I-215 to Canyon Lake - RC-1-34-2012 -- (RCFCWCD) 34 images
- 2013 Bautista Basin - RC-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Green Acres Dam - RC-1-7-2013 -- (RCFCWCD) 7 images
- 2013 Hemet Channel - RC-Hemet-1-24-2013 -- (RCFCWCD) 24 images
- 2013 Lakeview Dam - RC-Lakeview-1-4-2013 -- (RCFCWCD) 4 images
- 2013 Lamb Canyon Sanitary Landfill - RC-1-22-2013 -- (RCFCWCD) 22 images
- 2013 Lamb Canyon Sanitary Landfill Spot - RC-Spot-A-2013 -- (RCFCWCD) 1 image
- 2013 Park Hill Basin - RC-Park-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Romoland Main Drain Pipeline Line A, Stage 4 - RC-Romoland-1-47-2013 -- (RCFCWCD) 47 images
- 2013 San Jacinto Line E - RC-1-13-2013 -- (RCFCWCD) 13 images
- 2013 San Jacinto River I-215 to Canyon Lake - RC-1-34-2013 -- (RCFCWCD) 34 images
- 2013 San Jacinto River to Army Corp of Engineers to Bridge - RC-1-70-2013 -- (RCFCWCD) 70 images
- 2014 Lamb Canyon Sanitary Landfill - RC-1-18-2014 -- (RCFCWCD) 18 images
- 2014 Lamb Canyon Sanitary Landfill Spot- RC-A-2014 -- (RCFCWCD) 1 image
- 2014 Romoland Main Drain Pipeline Line A, Stage 4 - RC-1-47-2014 -- (RCFCWCD) 47 images
- 2014 San Jacinto River I-215 to Canyon Lake - RC-1-34-2014 -- (RCFCWCD) 34 images
- 2015 Hemet Channel - RC-1-24-2015 -- (RCFCWCD) 24 images
- 2015 San Jacinto Line E - RC-SanE-1-13-2015 -- (RCFCWCD) 13 images
- 2015 San Jacinto River Corp of Engineers to Bridge - RC-1-70-2015 -- (RCFCWCD) 70 images
- 2015 San Jacinto River I-215 to Canyon Lake - RC-1-34-2015 -- (RCFCWCD) 34 images
-
Lake Elsinore (2,877 images)
The Lake Elsinore area is in the southwestern section of Riverside County.
- 1948 Bundy Canyon Road - RC-94-100-1948 -- (RCFCWCD) 7 images
- 1948 Home Gardens to Railroad Canyon - RC-1-80-1948 -- (RCFCWCD) 75 images
- 1948 Railroad Canyon Southeast to County Line - RC-1-59-1948 -- (RCFCWCD) 44 images
- 1948 Temescal Creek Southeast to Lake Elsinore - RC-11-62-1948 -- (RCFCWCD) 32 images
- 1958 Elsinore - RC-41-51 -- (RCFCWCD) 11 images
- 1949 Elsinore Book 7A - RC-AXM-F-5F-12-174-1949 -- (RCFCWCD) 32 images
- 1949 Elsinore Book 7A - RC-AXM-F-9F-117-165-1949 -- (RCFCWCD) 23 images
- 1949 Elsinore Book 7A - RC-AXM-F-10F-49-218-1949 -- (RCFCWCD) 35 images
- 1949 Elsinore Book 13A - RC-AXM-F-11F-3-166-1949 -- (RCFCWCD) 13 images
- 1958 Lee Lake - RC-31-39 -- (RCFCWCD) 6 images
- 1960 Elsinore - RC-25-50 -- (RCFCWCD) 26 images
- 1961 Railroad Canyon - RC-1-3 -- (RCFCWCD) 2 images
- 1962 Elsinore - RC-8-16-1962 -- (RCFCWCD) 9 images
- 1963 Elsinore - RC-173-193-1963 -- (RCFCWCD) 21 images
- 1967 Elsinore Test Area - RC-3-8-8-7-1967 -- (RCFCWCD) 6 images
- 1967 Elsinore Test Area - RC-3-8-8-8-1967 -- (RCFCWCD) 6 images
- 1967 Railroad Canyon - RC-12-33-1967 -- (RCFCWCD) 22 images
- 1968 Elsinore-South Riverside Channel Convergence - RC-6-16-1968 -- (RCFCWCD) 11 images
- 1968 Elsinore-South Riverside Channel High Oblique - RC-17-1968 -- (RCFCWCD) 1 image
- 1968 La Cienega - RC-1-12-1968 -- (RCFCWCD) 12 images
- 1969 Catt Road Convergent - RC-1A-4-1969 -- (RCFCWCD) 5 images
- 1972 Lake Elsinore - RC-1-33-1972 -- (RCFCWCD) 23 images
- 1973 Massacre Canyon Borrow Site - RC-1-5-1973 -- (RCFCWCD) 5 images
- 1975 South Grand Avenue Channel - Lake Elsinore - RC-1-54-1975 -- (RCFCWCD) 30 images
- 1976 Elsinore Disposal - RC-Elsinore-3-5-1976 -- (RCFCWCD) 3 images
- 1976 South Grand Avenue Channel -RC-1-5-1976 -- (RCFCWCD) 5 images
- 1977 Elsinore Disposal - RC-Elsinore-1-3-1977 -- (RCFCWCD) 3 images
- 1979 Elsinore - RC-Elsinore-1-33-1979 -- (RCFCWCD) 33 images
- 1979 Elsinore Sanitary Landfill - RC-3-7-1979 -- (RCFCWCD) 5 images
- 1979 North Lake Elsinore - RC-LakeElsinore-1-5-1979 -- (RCFCWCD) 5 images
- 1979 South Grand Storm Drain - RC-1-3-1979 -- (RCFCWCD) 3 images
- 1979 South Grand Storm Drain - RC-1-10-1979 -- (RCFCWCD) 10 images
- 1979 Lakeland Village - RC-Lakeland-1-5-1979 -- (RCFCWCD) 5 images
- 1979 Lakeland Village - RC-Lakeland-1-10-1979 -- (RCFCWCD) 10 images
- 1979 Skylark Channel - RC-9-16-1979 -- (RCFCWCD) 8 images
- 1980 Lake Elsinore - RC-1-1980 -- (RCFCWCD) 1 image
- 1980 Lake Elsinore Flood - RC-1-245-1980 -- (RCFCWCD) 235 images
- 1980 Lake Elsinore Flood Obliques - RC-1-5-1980 -- (RCFCWCD) 4 images
- 1981 Leach Canyon Dam - RC-1-2-1981 -- (RCFCWCD) 2 images
- 1981 Leach Canyon Dam - RC-Leach-3-5-1981 -- (RCFCWCD) 3 images
- 1983 Elsinore - RC-1-6-1983 -- (RCFCWCD) 6 images
- 1983 Elsinore Sanitary Landfill - RC-Elsinore-1-6-1983 -- (RCFCWCD) 6 images
- 1983 Leach Canyon - RC-Leach-1-6-1983 -- (RCFCWCD) 6 images
- 1983 Sedco Line Drain - RC-1-6-1983 -- (RCFCWCD) 4 images
- 1983 Sedo Storm Drain - RC-1-16-1983 -- (RCFCWCD) 16 images
- 1983 Wasserman Canyon - RC-124-131-1983 -- (RCFCWCD) 8 images
- 1984 Lake Elsinore - Temescal Canyon - RC-1-99-1984 -- (RCFCWCD) 100 images
- 1984 Elsinore Sanitary Landfill - RC-Elsinore-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Leach Canyon Basin - RC-Leach-1-5-1984 -- (RCFCWCD) 5 images
- 1984 Leach Canyon Channel - RC-Leach-1-3-1984 -- (RCFCWCD) 3 images
- 1984 Sedco Ortho - RC-1-15-1984 -- (RCFCWCD) 15 images
- 1984 Sedco Ortho - RC-1-42-1984 -- (RCFCWCD) 40 images
- 1985 Leach Canyon Basin - RC-Leach2-1-5-1985 -- (RCFCWCD) 5 images
- 1985 Leach Canyon Channel - RC-Leach-1-3-1985 -- (RCFCWCD) 3 images
- 1985 Ocotillo Storm Drain - RC-1-11-1985 -- (RCFCWCD) 11 images
- 1985 Ocotillo Storm Drain - RC-1-33-1985 -- (RCFCWCD) 33 images
- 1985 Ocotillo Storm Drain - RC-Ocotillo-1-4-1985 -- (RCFCWCD) 4 images
- 1985 South Riverside Channel - RC-South-1-3-1985 -- (RCFCWCD) 3 images
- 1986 Arroyo del Toro - RC-1-10-1986 -- (RCFCWCD) 10 images
- 1986 Lake Elsinore Outlet Channel - RC-1-68-1986 -- (RCFCWCD) 67 images
- 1986 Four Corners Storm Drain - RC-1-32-1986 -- (RCFCWCD) 32 image
- 1986 Wasson Canyon Channel - RC-1-9-1986 -- (RCFCWCD) 9 images
- 1987 Leach Canyon Basin - RC-Leach-1-6-1987 -- (RCFCWCD) 6 images
- 1987 Leach Canyon Channel - RC-Leach2-1-3-1987 -- (RCFCWCD) 3 images
- 1987 South Riverside Channel - RC-Riverside-1-4-1987 -- (RCFCWCD) 4 images
- 1988 Leach Canyon - RC-Leach-1-7-1988 -- (RCFCWCD) 7 images
- 1988 Leach Canyon Channel - RC-Leach2-1-4-1988 -- (RCFCWCD) 4 images
- 1988 South Riverside Channel - RC-South-1-3-1988 -- (RCFCWCD) 3 images
- 1988 Vista Way - RC-Vista-1-9-1988 -- (RCFCWCD) 9 images
- 1989 Elsinore Outlet CH - RC-1-20-1989 -- (RCFCWCD) 20 images
- 1989 Lamb Canyon - RC-Lamb-1-1989 -- (RCFCWCD) 1 images
- 1989 Lamb Canyon Sanitary Landfill - RC-Lamb2-1-6-1989 -- (RCFCWCD) 6 images
- 1989 Leach Canyon Basin - RC-Leach-1-5-1989 -- (RCFCWCD) 5 images
- 1989 Leach Canyon Channel - RC-Leach-1-3-1989 -- (RCFCWCD) 3 images
- 1989 Mission Trail - RC-1-8-1989 -- (RCFCWCD) 8 images
- 1989 Ortega Channel - RC-Ortega-1-3-1989 -- (RCFCWCD) 3 images
- 1989 South Riverside Channel - RC-South-1-4-1989 -- (RCFCWCD) 4 images
- 1990 Leach Canyon Basin - RC-Leach-1-6-1990 -- (RCFCWCD) 6 images
- 1990 Railroad Canyon Ortho - RC-1-26-1990 -- (RCFCWCD) 26 images
- 1990 Railroad Canyon Ortho - RC-1-76-1990 -- (RCFCWCD) 76 images
- 1990 Railroad Canyon Ortho - RC-Railroad-1-13-1990 -- (RCFCWCD) 13 images
- 1991 Elsinore - RC-1-41-1991 -- (RCFCWCD) 41 images
- 1991 Elsinore Sanitary Landfill - RC-Elsinore-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Lakeland Village Channel - RC-Lakeland-1-8-1991 -- (RCFCWCD) 8 images
- 1991 Leach Canyon - RC-Leach-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Leach Canyon Basin - RC-Leach-1-5-1991 -- (RCFCWCD) 5 images
- 1991 Limestreet Channel - RC-1-9-1991 -- (RCFCWCD) 9 images
- 1991 Ortega Channel - RC-Ortega-1-9-1991 -- (RCFCWCD) 9 images
- 1991 South Riverside Channel - RC-South2-1-4-1991 -- (RCFCWCD) 4 images
- 1992 Leach Canyon Basin - RC-1-4-1992 -- (RCFCWCD) 4 images
- 1993 Lake Elsinore Outlet - RC-Elsinore-1-9-1993 -- (RCFCWCD) 9 images
- 1993 Lake Elsinore Spot - RC-Spot-1-1993 -- (RCFCWCD) 1 image
- 1993 Lime Street Channel - RC-Lime-1-4-1993 -- (RCFCWCD) 4 images
- 1994 Elsinore Sanitary Landfill - RC-Elsinore2-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Elsinore Spot - RC-Elsinore-A-1994 -- (RCFCWCD) 1 image
- 1994 Lime Street Channel - RC-Lime-1-4-1994 -- (RCFCWCD) 4 images
- 1994 South Riverside Channel - RC-South-1-4-1994 -- (RCFCWCD) 4 images
- 1995 Elsinore Sanitary Landfill Spot - RC-Elsinore-A-1995 -- (RCFCWCD) 1 image
- 1995 Elsinore Sanitary Landfill - RC-Elsinore2-1-4-1995 -- (RCFCWCD) 4 images
- 1995 Lake Elsinore - RC-1-34-1995 -- (RCFCWCD) 34 images
- 1995 Lake Elsinore Outlet - RC-1-27-1995 -- (RCFCWCD) 27 images
- 1995 Leach Canyon Dam - RC-Leach-1-7-1995 -- (RCFCWCD) 7 images
- 1995 Wasson Canyon Debris Basin - RC-Wasson-1-3-1995 -- (RCFCWCD) 3 images
- 1995 West Elsinore Line A - RC-West-1-4-1995 -- (RCFCWCD) 4 images
- 1996 Elsinore Sanitary Landfill - RC-Elsinore-1-4-1996 -- (RCFCWCD) 4 images
- 1996 Leach Canyon Basin - RC-Leach-1-6-1966 -- (RCFCWCD) 6 images
- 1996 Leach Canyon Channel - RC-Leach2-1-3-1996 -- (RCFCWCD) 3 images
- 1996 Lime Street Channel - RC-Lime-1-4-1996 -- (RCFCWCD) 4 images
- 1996 Ortega Channel - RC-2-14-1996 -- (RCFCWCD) 13 images
- 1996 South Riverside Channel - RC-South-1-5-1996 -- (RCFCWCD) 5 images
- 1997 Lakeland Village - RC-1-2-1997 -- (RCFCWCD) 2 images
- 1997 Leach Canyon Channel - RC-Leach-1-3-1997 -- (RCFCWCD) 3 images
- 1997 Lime Street Channel - RC-Lime-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Ortega Channel - RC-Ortega-1-3-1997 -- (RCFCWCD) 3 images
- 1997 South Riverside Channel - RC-South-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Wasson Basin - RC-Wasson-1-3-1997 -- (RCFCWCD) 3 images
- 1998 Lake Elsinore Outlet - RC-1-33-1998 -- (RCFCWCD) 35 images
- 1998 Lakeland Village Channel - RC-Lakeland-1-2-1998 -- (RCFCWCD) 2 images
- 1998 Leach Canyon - RC-Leach-1-6-1998 -- (RCFCWCD) 6 images
- 1998 Leach Canyon Channel - RC-Leach2-1-3-1998 -- (RCFCWCD) 3 images
- 1998 Lime Street Channel - RC-Lime-1-4-1998 -- (RCFCWCD) 4 images
- 1998 Ortega Channel - RC-Ortega-1-2-1998 -- (RCFCWCD) 2 images
- 1998 Ortega Channel Debris Basin - RC-1-2-1998 -- (RCFCWCD) 2 images
- 1998 South Riverside Channel - RC-South-1-3-1998 -- (RCFCWCD) 3 images
- 1998 Wasson Basin - RC-Wasson-1-3-1998 -- (RCFCWCD) 3 images
- 1999 Elsinore Outlet Channel - RC-1-31-1999 -- (RCFCWCD) 31 images
- 1999 Lakeland Village Channel - RC-Lakeland-1-2-1999 -- (RCFCWCD) 2 images
- 1999 Leach Canyon - RC-Leach-1-3-1999 -- (RCFCWCD) 3 images
- 1999 Leach Canyon Dam - RC-Leach2-1-6-1999 -- (RCFCWCD) 6 images
- 1999 Lime Street Channel - RC-Lime-1-4-1999 -- (RCFCWCD) 4 images
- 1999 Ortega Channel - RC-Ortega-1-3-1999 -- (RCFCWCD) 3 images
- 1999 South Riverside Channel - RC-South-1-4-1999 -- (RCFCWCD) 4 images
- 1999 Wasson Canyon Basin - RC-Wasson-1-3-1999 -- (RCFCWCD) 3 images
- 2000 Elsinore Outlet Channel - RC-1-31-2000 -- (RCFCWCD) 31 images
- 2000 Elsinore Sanitary Landfill - RC-Elsinore2-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Elsinore Sanitary Landfill Spot - RC-Elsinore-A-2000 -- (RCFCWCD) 1 image
- 2000 Lakeland Village Channel - RC-Lakeland-1-2-2000 -- (RCFCWCD) 2 images
- 2000 Leach Canyon Channel - RC-Leach-1-6-2000 -- (RCFCWCD) 4 images
- 2000 Lime Street Channel - RC-Lime-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Ortega Channel - RC-Ortega-1-4-2000 -- (RCFCWCD) 4 images
- 2000 South Riverside Channel - RC-South-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Wasson Basin - RC-Wasson-1-3-2000 -- (RCFCWCD) 3 images
- 2001 Elsinore Outlet - RC-1-6-2001 -- (RCFCWCD) 6 images
- 2001 Elsinore Outlet - RC-1-28-2001 -- (RCFCWCD) 28 images
- 2001 Lakeland Village Channel - RC-Lakeland-1-2-2001 -- (RCFCWCD) 2 images
- 2001 Leach Canyon Channel - RC-Leach-1-3-2001 -- (RCFCWCD) 3 images
- 2001 Leach Canyon Dam - RC-Leach2-1-6-2001 -- (RCFCWCD) 6 images
- 2001 Lime Street Channel - RC-Lime-1-4-2001 -- (RCFCWCD) 4 images
- 2001 South Riverside Channel - RC-South-1-4-2001 -- (RCFCWCD) 4 images
- 2001 Wasson Basin - RC-Wasson-1-3-2001 -- (RCFCWCD) 3 images
- 2002 Elsinore East Ortho - RC-1-48-2002 -- (RCFCWCD) 48 images
- 2002 Elsinore West Oertho - RC-1-70-2002 -- (RCFCWCD) 70 images
- 2002 Elsinore West Ortho A & B - RC-A-B-2002 -- (RCFCWCD) 2 images
- 2002 Lakeland Village Channbel - RC-Lakeland-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Leach Canyon Channel - RC-Leach-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Leach Canyon Channel - RC-Leach2-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Lime Street Channel - RC-Lime-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Ortega Channel - RC-Ortega-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Prenda Dam - RC-Prenda-1-3-2002 -- (RCFCWCD) 3 images
- 2002 South Riverside Channel - RC-South-1-3-2002 -- (RCFCWCD) 3 images
- 2003 Elsinore Outlet Channel - RC-1-19-2003 -- (RCFCWCD) 19 images
- 2003 Elsinore Outlet Channel - RC-Elsinore-1-19-2003 -- (RCFCWCD) 19 images
- 2003 Lakeland Village - RC-Lakeland-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Leach Canyon Channel - RC-Leach-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Leach Canyon Dam - RC-Leach2-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Lime Street Channel - RC-Lime-1-3-2003 -- (RCFCWCD) 3 images
- 2003 McVicker Canyon Basin - RC-McVicker-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Ortega Basin - RC-Ortega-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Ortega Channel - RC-Ortega-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Sedco Basin - RC-Sedco-1-2-2003 -- (RCFCWCD) 2 images
- 2004 Wasson Basin - RC-Wasson-1-3-2004 -- (RCFCWCD) 3 images
- 2003 South Riverside Channel - RC-South-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Wasson Basin - RC-Wasson-1-3-2003 -- (RCFCWCD) 3 images
- 2004 Edom Hill Sanitary Landfill - RC-1-23-2004 -- (RCFCWCD) 23 images
- 2004 Elsinore Outlet Channel - RC-Elsinore-1-19-2004 -- (RCFCWCD) 19 images
- 2004 Lakeland Village - RC-Lakeland-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Leach Canyon Channel - RC-Leach2-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Leach Canyon Dam - RC-Leach-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Lime Street Channel - RC-Lime-1-3-2004 -- (RCFCWCD) 3 images
- 2004 McVicker Canyon Basin - RC-McVicker-1-7-2004 -- (RCFCWCD) 7 images
- 2004 Ortego Basin - RC-Ortego2-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Ortega Basin - RC-Ortega-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Ortega Channel - RC-Ortega-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Sedco Basin - RC-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Sedco Basin - RC-Sedco-1-2-2004 -- (RCFCWCD) 2 images
- 2004 South Riverside Channel - RC-South-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Wasson Basin - RC-Wasson-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Wasson Basin - RC-Wasson-1-3-2004 -- (RCFCWCD) 3 images
- 2005 Arroyo Del Toro Canyon - RC-Arroyo-1-2005 -- (RCFCWCD) 1 image
- 2005 Arroyo Del Toro - RC-Arroyo2-1-8-2005 -- (RCFCWCD) 8 images
- 2005 Elsinore Outlet Channel - RC-Elsinore7-1-20-2005 -- (RCFCWCD) 20 images
- 2005 Gunnerson Pond - RC-1-4-2005 -- (RCFCWCD) 4 images
- 2005 Gunnerson Pond - RC-Pond-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Lakeland Village - RC-Lakeland7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Lakeland Village & Sedco - RC-1-57-2005 -- (RCFCWCD) 57 images
- 2005 Leach Canyon Channel - RC-Leach8-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Leach Canyon Dam - RC-Leach7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Leach Canyon Dam - RC-Leach-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Lime Street Channel - RC-Lime7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 McVicker Canyon Basin - RC-McVicker2-1-6-2005 -- (RCFCWCD) 7 images
- 2005 McVicker Canyon Basin - RC-McVicker-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Ortega Basin - RC-Ortega8-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Ortega Channel - RC-Ortega7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 South Riverside Channel - RC-Riverside7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Wasson Basin - RC-Wasson7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Wasson Canyon - RC-Wasson-1-2005 -- (RCFCWCD) 1 image
- 2006 Elsinore Outlet Channel - RC-1-19-2006 -- (RCFCWCD) 19 images
- 2006 Lake Elsinore - RC-30-54-2006 -- (RCFCWCD) 25 images
- 2006 Lake Elsinore - RC-55-64-2006 -- (RCFCWCD) 10 images
- 2006 Lake Elsinore - RC-65A-74-2006 -- (RCFCWCD) 11 images
- 2006 Lake Elsinore - RC-94-97-2006 -- (RCFCWCD) 4 images
- 2006 Lake Elsinore - RC-Elsinore2-1-18-2006 -- (RCFCWCD) 18 images
- 2006 Lakeland Village Channel - RC-Lakeland-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Leach Canyon Channel - RC-Leach7-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Leach Canyon Dam - RC-Leach6-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Lime Street Channel - RC-Lime-1-3-2006 -- (RCFCWCD) 3 images
- 2006 McVicker Canyon Basin - RC-McVicker7-1-7-2006 -- (RCFCWCD) 7 images
- 2006 Ortega Basin - RC-Ortega-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Ortega Channel - RC-Ortega6-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Railroad Canyon - RC-1-55-2006 -- (RCFCWCD) 55 images
- 2006 Sedco Basin - RC-Sedco-1-2-2006 -- (RCFCWCD) 2 images
- 2006 South Riverside Channel - RC-South2-1-3-2006 -- (RCFCWCD) 3 images
- 2007 Elsinore Outlet Channel - RC-1-19-2007 -- (RCFCWCD) 19 images
- 2007 Lakeland Village Channel - RC-Lakeland-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Leach Canyon Channel - RC-Leach-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Leach Canyon Dam - RC-Leach2-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Lime Street Channel - RC-Lime-1-3-2007 -- (RCFCWCD) 3 images
- 2007 McVicker Canyon Basin - RC-McVicker-1-6-2007 -- (RCFCWCD) 6 images
- 2007 Ortega Channel - RC-Ortega-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Ortega Basin - RC-Ortega2-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Sedco Basin - RC-Sedco-1-2-2007 -- (RCFCWCD) 2 images
- 2007 South Riverside Channel - RC-South-1-4-2007 -- (RCFCWCD) 4 images
- 2008 Lakeland Village Orange Street - RC-Lakeland7-1-3-2008 -- (RCFCWCD) 3 images
- 2007 Wasson Basin - RC-Wasson-1-3-2007 -- (RCFCWCD) 3 images
- 2009 Elsinore Outlet Channel - RC-1-19-2009 -- (RCFCWCD) 19 images
- 2009 Elsinore Outlet - RC-Elsinore7-1-19-2009 -- (RCFCWCD) 19 images
- 2009 Gunnerson Pond Obliques - RC-Gunnerson-1-2009 -- (RCFCWCD) 1 image
- 2009 Lakeland Village Channel - RC-Lakeland-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Lakeland Village Channel - RC-Lakeland7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Leach Canyon Channel - RC-Leach7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Leach Canyon Channel - RC-Leach-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Leach Canyon Dam - RC-Leach8-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Leach Canyon Dam - RC-Leach2-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Lime Street Channel - RC-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Lime Street Channel - RC-Lime-1-3-2009 -- (RCFCWCD) 3 images
- 2009 McVicker Canyon Basin - RC-McVicker-1-6-2009 -- (RCFCWCD) 6 images
- 2009 McVicker Canyon Basin - RC-1-6-2009 -- (RCFCWCD) 6 images
- 2009 Ortega Basin - RC-Ortega7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Ortega Basin - RC-Ortega8-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Ortega Channel - RC-Ortega7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Ortega Channel - RC-Ortega-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Sedco Basin - RC-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Sedco Basin - RC-Sedco-1-2-2009 -- (RCFCWCD) 2 images
- 2009 South Riverside Channel - RC-South8-1-3-2009 -- (RCFCWCD) 3 images
- 2009 South Riverside Channel - RC-South-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Wasson Basin - RC-Wasson7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Wasson Basin - RC-Wasson-1-3-2009 -- (RCFCWCD) 3 images
- 2010 Gunnerson Pond - RC-1-10-2010 -- (RCFCWCD) 10 images
- 2011 Elsinore Outlet Channel - RC-1-19-2011 -- (RCFCWCD) 19 images
- 2011 Lakeland Village Channel - RC-Lakeland-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Leach Canyon - RC-Leach-1-3-2011 -- (RCFCWCD) 4 images
- 2011 Leach Canyon - RC-Leach2-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Lime Street Channel - RC-Lime-1-2-2011 -- (RCFCWCD) 3 images
- 2011 McVicker Canyon Basin - RC-McVicker-1-6-2011 -- (RCFCWCD) 6 images
- 2011 Ortega Basin - RC-Ortega-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Ortega Channel - RC-Ortega2-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Sedco Basin - RC-Sedco-1-2-2011 -- (RCFCWCD) 2 images
- 2011 South Riverside Channel - RC-South-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Wasson Basin - RC-Wasson-1-3-2011 -- (RCFCWCD) 3 images
- 2012 Elsinore Outlet Channel - RC-1-19-2012 -- (RCFCWCD) 19 images
- 2012 Lakeland Village Channel - RC-Lakeland-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Leach Canyon Dam - RC-Leach-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Leach Canyon Channel - RC-Leach2-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Lime Street Channel - RC-Lime-1-3-2012 -- (RCFCWCD) 3 images
- 2012 McVicker Canyon Basin - RC-McVicker-1-6-2012 -- (RCFCWCD) 6 images
- 2012 Ortega Basin - RC-Ortega-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Ortega Channel - RC-Ortega2-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Sedco Basin - RC-Sedco-1-2-2012 -- (RCFCWCD) 2 images
- 2012 South Riverside Channel - RC-South-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Wasson Basin - RC-Wasson-1-3-2012 -- (RCFCWCD) 3 images
- 2013 Elsinore Outlet Channel - RC-1-19-2013 -- (RCFCWCD) 19 images
- 2013 Lakeland Village Adelfa Street Channel - RC-1-30-2013 -- (RCFCWCD) 30 images
- 2013 Lakeland Village Channel - RC-Lakeland-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Leach Canyon Channel - RC-Leach-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Lime Street Channel - RC-Lime-1-3-2013 -- (RCFCWCD) 3 images
- 2013 McVicker Canyon Basin - RC-McVicker-1-6-2013 -- (RCFCWCD) 6 images
- 2013 Ortega Channel - RC-Ortega-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Ortega Basin - RC-Ortega2-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Sedco Basin - RC-Sedco-1-2-2013 -- (RCFCWCD) 2 images
- 2013 South Riverside Channel - RC-South-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Wasson Basin - RC-Wasson-1-3-2013 -- (RCFCWCD) 3 images
- 2015 Elsinore Outlet Channel - RC-1-19-2015 -- (RCFCWCD) 19 images
- 2015 Lakeland Village Channel - RC-Lakeland-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Leach Canyon Channel - RC-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Leach Canyon Dam - RC-Leach-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Lime Street Channel - RC-Lime-1-3-2015 -- (RCFCWCD) 3 images
- 2015 McVicker Canyon Basin - RC-McVicker-1-6-2015 -- (RCFCWCD) 6 images
- 2015 Ortega Basin - RC-Ortega2-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Ortega Channel - RC-Ortega-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Prenda Dam - RC-Prenda-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Sedco Basin - RC-Sedco-1-2-2015 -- (RCFCWCD) 2 images
- 2015 South Riverside Channel - RC-South-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Wasson Basin - RC-Wasson-1-3-2015 -- (RCFCWCD) 3 images
-
Moreno Valley (7,982 images)
The Moreno Valley area is east of the City of Riverside and invludes March Air Force Base, Perris, Sun City, Lakeview, and Winchester.
- 1948 Bautista Creek - RC-24-72-1948 -- (RCFCWCD) 47 images
- 1948 Bautista Creek - RC-164-240-1948 -- (RCFCWCD) 76 images
- 1948 March Air Force Base - RC-1-34-1948 -- (RCFCWCD) 33 images
- 1948 March Air Force Base to Corona - RC-5-144-1948 -- (RCFCWCD) 117 images
- 1948 March Air Force Base to Hemet - RC-8-209-1948 -- (RCFCWCD) 138 images
- 1949 Bautista Creek Obliques - RC-32-35-1949 -- (RCFCWCD) 4 images
- 1949 Perris Book 4-A - RC-AXM-F-11F-62-162-1949 -- (RCFCWCD) 15 images
- 1949 Perris Book 4-A - RC-AXM-F-12F-46-156-1949 -- (RCFCWCD) 18 images
- 1952 Pigeon Pass to Lake Matthews - RC-1-43-1952 -- (RCFCWCD) 43 images
- 1952 Pigeon Pass to Lake Matthews - RC-2-21-42-1952 -- (RCFCWCD) 22 images
- 1959 San Jacinto River & Perris - RC-20-36 -- (RCFCWCD) 36 images
- 1959 Sunnymead - RC-25-33 -- (RCFCWCD) 9 images
- 1959 Sunnymead - RC-34-40 -- (RCFCWCD) 7 images
- 1959 Sunnymead/Moreno Valley - RC-14-38 -- (RCFCWCD) 10 images
- 1960 March Field - RC-20-24 -- (RCFCWCD) 5 images
- 1960 Moreno Valley - RC-13-19 -- (RCFCWCD) 7 images
- 1962 East Riverside/Sunnymead - RC-43-53-1962 -- (RCFCWCD) 11 images
- 1962 East Riverside/Sunnymead - RC-54-84-1962 -- (RCFCWCD) 32 images
- 1962 Sunnymead - RC-82-107-1962 -- (RCFCWCD) 26 images
- 1964 Perris Valley - RC-57-108-1964 -- (RCFCWCD) 52 images
- 1965 North Sunnymead - RC-1-28-1965 -- (RCFCWCD) 28 images
- 1966 Alessandro Blvd - RC-1A-6A-1966 -- (RCFCWCD) 5 images
- 1966 Lakeview Part 1 - RC-39-76-1966 -- (RCFCWCD) 38 images
- 1966 Lakeview Part 2 - RC-49-95-1966 -- (RCFCWCD) 47 images
- 1967 Romoland - RC-18-51-1967 -- (RCFCWCD) 33 images
- 1967 Sun City - RC-28-73-1967 -- (RCFCWCD) 46 images
- 1968 Good Hope - RC-16-42-1968 -- (RCFCWCD) 27 images
- 1969 Alberhill - RC-18-49-1969 -- (RCFCWCD) 30 images
- 1969 March Field -RC-18-57-1969 -- (RCFCWCD) 41 images
- 1969 Perris - Route 81 - RC-1-18-1969 -- (RCFCWCD) 18 images
- 1969 Perris - Cajalco Road - RC-19-45-1969 -- (RCFCWCD) 27 images
- 1969 San Jacinto River (Lakeview) - RC-101-179-1969 -- (RCFCWCD) 80 images
- 1970 Gavilan Hills - RC-27-74-1970 -- (RCFCWCD) 48 images
- 1970 Perris Boulevard - RC-10-15-1970 -- (RCFCWCD) 6 images
- 1971 Alessandro - RC-4-10-1971 -- (RCFCWCD) 7 images
- 1971 Alessandro & Trautwein - RC-3-1971 -- (RCFCWCD) 1 image
- 1971 Lamb Canyon Dumpsite - RC-13-27-1971 -- (RCFCWCD) 15 images
- 1971 Menifee & St. Highway 74 - RC-11-1971 -- (RCFCWCD) 1 image
- 1971 Perris - RC-1-5-1971 -- (RCFCWCD) 1 images
- 1971 Perris - RC-1-6-1971 -- (RCFCWCD) 6 images
- 1971 Pettit Street/Sunnymead - RC-16-50-1971 -- (RCFCWCD) 35 images
- 1971 Pigeon Pass - RC-Pigeon-1-9-1971 -- (RCFCWCD) 9 images
- 1971 Ramona Expressway & Sanders - RC-12-1971 -- (RCFCWCD) 1 image
- 1972 Bautista Creek - RC-1-14-1972 -- (RCFCWCD) 14 images
- 1973 Alessandro Blvd. - RC-56-77-1973 -- (RCFCWCD) 21 images
- 1973 Cajalco Road - RC-1-42-1973 -- (RCFCWCD) 42 images
- 1973 Kitching Street - RC-40-55-1973 -- (RCFCWCD) 16 images
- 1973 Perris Boulevard - RC-1-9-1973 -- (RCFCWCD) 9 images
- 1974 Perris Boulevard - RC-1-35-1974 -- (RCFCWCD) 35 images
- 1975 Mead Valley - RC-Mead-1-4-1975 -- (RCFCWCD) 4 images
- 1976 Lakeview - RC-1-28-1976 -- (RCFCWCD) 19 images
- 1976 Moreno - RC-15-26-1976 -- (RCFCWCD) 12 images
- 1976 Perris City Drains - RC-1-11-1976 -- (RCFCWCD) 11 images
- 1976 Perris City Drains - RC-Perris-1-8-1976 -- (RCFCWCD) 8 images
- 1976 Perris Drains - RC-13-27-1976 -- (RCFCWCD) 15 images
- 1976 Perris Valley - RC-1-60-1976 -- (RCFCWCD) 60 images
- 1977 Hemet - RC-1-60-1977 -- (RCFCWCD) 59 images
- 1977 Hemet High Oblique - RC-82-84-1977 -- (RCFCWCD) 2 images
- 1977 Little Lake - RC-Little-1-6-1977 -- (RCFCWCD) 6 images
- 1977 Little Lake Mosaic -RC-1-80-1976 -- (RCFCWCD) 70 images
- 1977 Salt Creek - RC-1-24-1977 -- (RCFCWCD) 18 images
- 1977 Salt Creek - RC-59-140-1977 -- (RCFCWCD) 82 images
- 1977 Salt Creek Obliques - RC-25-28-1977 -- (RCFCWCD) 4 images
- 1977 Sunnymead - RC-1-45-1977 -- (RCFCWCD) 45 images
- 1978 Park Hill Basin - RC-1-53-1978 -- (RCFCWCD) 53 images
- 1978 Park Hill Basin - RC-2-36-1978 -- (RCFCWCD) 35 images
- 1978 Park Hill Basin - RC-7-15-1978 -- (RCFCWCD) 8 images
- 1979 Little Lake Basin - RC-1-33-1979 -- (RCFCWCD) 33 images
- 1979 Little Lake Basin - RC-5-10-1979 -- (RCFCWCD) 6 images
- 1979 Meridian Street Channel - RC-1-13-1979 -- (RCFCWCD) 13 images
- 1979 Meridian Street Channel - RC-1-23-1979 -- (RCFCWCD) 23 images
- 1979 North Perris Third Street Basin - RC-1-27-1979 -- (RCFCWCD) 27 images
- 1979 North Perris Third Street Basin - RC-Perris-2-3-5-1979 -- (RCFCWCD) 3 images
- 1979 Sunnymead Ramp 9 - RC-1-55-1979 -- (RCFCWCD) 55 images
- 1980 Pidgeon Pass Dam - RC-1-8-1980 -- (RCFCWCD) 8 images
- 1982 Sunnymead - RC-1-14-1982 -- (RCFCWCD) 14 images
- 1983 Bautistia - RC-Bautista-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Metz - RC-Metz-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Parkhill - RC-Parkhill-1-3-1983 -- (RCFCWCD) 3 images
- 1983 Perris Third Street - RC-Perris-1-3-1983 -- (RCFCWCD) 3 images
- 1983 Pigeon Pass Basin - RC-Pigeon-1-6-1983 -- (RCFCWCD) 6 images
- 1984 Lakeview Dam - RC-Lakeview-1-12-1984 -- (RCFCWCD) 12 images
- 1984 Lakeview Dam Outlet Channel - RC-1-25-1984 -- (RCFCWCD) 25 images
- 1984 Metz Basin - RC-Metz-1-5-1984 -- (RCFCWCD) 6 images
- 1984 Perris Third Street Basin - RC-Perris-1-3-1984 -- (RCFCWCD) 3 images
- 1984 Pigeon Pass Basin - RC-Pigeon-1-7-1984 -- (RCFCWCD) 7 images
- 1985 Bautista Creek - RC-1-45-1985 -- (RCFCWCD) 42 images
- 1985 De Anza Cycle Park - RC-1-105-1985 -- (RCFCWCD) 80 images
- 1985 Little Lake Basin - RC-Little-1-2-1985 -- (RCFCWCD) 2 images
- 1985 Metz Basin - RC-Metz-1-5-1985 -- (RCFCWCD) 5 images
- 1985 Parkhill Basin - RC-Parkhill-1-2-1985 -- (RCFCWCD) 2 images
- 1985 Perris Third Street Basin - RC-Perris-1-3-1985 -- (RCFCWCD) 3 images
- 1985 Perris Valley - RC-1-183-1985 -- (RCFCWCD) 181 images
- 1985 Pigeon Pass - RC-Pigeon-1-7-1985 -- (RCFCWCD) 5 images
- 1985 Sunnymead Line H-7 - RC-2-4-1985 -- (RCFCWCD) 3 images
- 1985 Sunnymead Line M-11Stage 1 - RC-1-13-1985 -- (RCFCWCD) 13 images
- 1985 Sunnymead Line P - RC-Sunnymead-1-17-1985 -- (RCFCWCD) 17 images
- 1986 Bautista Creek Channel - RC-1-41-1986 -- (RCFCWCD) 40 images
- 1986 Bautista Creek - RC-Bautista-1-10-1986 -- (RCFCWCD) 10 images
- 1987 Good Hope Channel - RC-1-6-1987 -- (RCFCWCD) 6 images
- 1986 Ironwood Avenue - RC-1-11-1986 -- (RCFCWCD) 11 images
- 1987 Moreno Vellay Line F - RC-1-27-1987 -- (RCFCWCD) 27 images
- 1986 Ramona Expressway 86 - RC-1-25-1986 -- (RCFCWCD) 25 images
- 1986 Sunnymead Line 13-B - RC-1-22-1986 -- (RCFCWCD) 22 images
- 1987 Heacock Channel - RC-Heacock-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Pigeon Pass Basin - RC-Pigeon-1-6-1987 -- (RCFCWCD) 6 images
- 1987 Pigeon Pass Dam - RC-Pigeon-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Sunnymead Line H-12 - RC-1-11-1987 -- (RCFCWCD) 11 images
- 1987 Sunnymead Line P - RC-1-17-1987 -- (RCFCWCD) 16 images
- 1988 Bautista Basin - RC-Bautista-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Bautista Channel - RC-1-39-1988 -- (RCFCWCD) 39 images
- 1988 Juniper Flats Excavation - RC-Juniper-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Lakeview Dam - RC-Lakeview-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Moreno Line F-2 - RC-1-16-1988 -- (RCFCWCD) 16 images
- 1988 Perris Valley Line J - RC-1-8-1988 -- (RCFCWCD) 8 images
- 1988 San Jacinto Phase III - RC-1-27-1988 -- (RCFCWCD) 27 images
- 1988 San Jacinto River - RC-1-31-1988 -- (RCFCWCD) 31 images
- 1988 Sunnymead MDP-B6 - RC-Sunnymead2-1-2-1988 -- (RCFCWCD) 2 images
- 1989 Heacock Channel - RC-1-12-1989 -- (RCFCWCD) 12 images
- 1989 Heacock Lateral A - RC-1-14-1989 -- (RCFCWCD) 14 images
- 1989 Moreno F-2 - RC-Moreno-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Moreno Line K-1 - RC-1-1-1-3-1989 -- (RCFCWCD) 6 images
- 1989 Perris Valley Obliques - RC-Perris2-1-4-1989 -- (RCFCWCD) 6 images
- 1989 Romoland Channel - RC-Romoland-1-8-1989 -- (RCFCWCD) 8 images
- 1989 San Jacinto - RC-1-53-1989 -- (RCFCWCD) 53 images
- 1989 Sunnymead Line H-1 - RC-1-33-1989 -- (RCFCWCD) 33 images
- 1990 Hospital MV-UCR - RC-Hospital-1-4-1990 -- (RCFCWCD) 4 images
- 1990 Nason Strret/Riverside County Hospital - RC-Nason-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Old Bautista Wash - RC-1-15-1990 -- (RCFCWCD) 15 images
- 1990 Ramona Expressway - RC-Ramona-1-14-1990 -- (RCFCWCD) 14 images
- 1991 Canyon Springs Retention Basin - RC-Canyon-1-2-1991 -- (RCFCWCD) 2 images
- 1991 County Hospital Cactus - RC-County-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Esplanade & Sanderson - RC-1-6-1991 -- (RCFCWCD) 6 images
- 1991 Lamb Canyon Sanitary Landfill - RC-Lamb-1-6-1991 -- (RCFCWCD) 6 images
- 1991 LaSalle Basin - RC-LaSalle-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Bautista Basin - RC-Bautista-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Lakeview Ortho Phase 2 - RC-1-38-1991 -- (RCFCWCD) 38 images
- 1991 Little Lake Basin - RC-Little-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Metz Basin - RC-Metz-1-5-1991 -- (RCFCWCD) 5 images
- 1991 Perris Boulevard - RC-Perris-1-1991 -- (RCFCWCD) 19 images
- 1991 Pigeon Pass Basin - RC-Pigeon-1-6-1991 -- (RCFCWCD) 6 images
- 1992 Bautista Basin - RC-Bautista-1-3-1992 -- (RCFCWCD) 3 images
- 1992 Buena Vista Basin - RC-Buena-1-3-1992 -- (RCFCWCD) 3 images
- 1992 Canyon Springs Retention Basin - RC-Canyon-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Heacock/Cactus - RC-Heacock-1-10-1992 -- (RCFCWCD) 10 images
- 1992 Lassalle Basin - RC-Lassalle-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Little Lake Basin - RC-Little-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Little San Gorgonio Spreading Grounds - RC-LIttle2-1-5-1992 -- (RCFCWCD) 5 images
- 1992 Moreno F-2 - RC-1-14-1992 -- (RCFCWCD) 14 images
- 1992 Moreno Valley - WO-12-92-1992 -- (Imbiorski) 3 images
- 1992 Park Hill Basin - RC-Park-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Perris Third Street Basin - RC-Perris-1-3-1992 -- (RCFCWCD) 3 images
- 1992 San Jacinto River - RC-1-43-1992 -- (RCFCWCD) 43 images
- 1992 Sunnymead Line K-1 - RC-1-6-1992 -- (RCFCWCD) 6 images
- 1992 Sunnymead M-11 Stage 2 - RC-1-8-1992 -- (RCFCWCD) 8 images
- 1992 Sunnymead M-16 - RC-1-10-1992 -- (RCFCWCD) 10 images
- 1993 Bradley Road Storm Drain - RC-1-5-1993 -- (RCFCWCD) 5 images
- 1993 Perris Valley - RC-1-255-1993 -- (RCFCWCD) 252 images
- 1993 Perris Valley - RC-153-255-1993 -- (RCFCWCD) 103 images
- 1993 Perris Valley Line J - RC-1-10-1993 -- (RCFCWCD) 10 images
- 1993 Salt Creek - RC-1-156-1993 -- (RCFCWCD) 155 images
- 1993 Salt Creek - RC-156-177-1993 -- (RCFCWCD) 22 images
- 1993 San Jacinto Line E - RC-Line-E-1-26-1993 -- (RCFCWCD) 27 images
- 1993 San Jacinto River - RC-1-101-1993 -- (RCFCWCD) 101 images
- 1993 San Jacinto River - RC-102-113-1993 -- (RCFCWCD) 12 images
- 1993 San Jacinto River - RC-1-93-1993 -- (RCFCWCD) 93 images
- 1993 Sunnymead MDP Line F - RC-Sunnymnead-1-6-1993 -- (RCFCWCD) 6 images
- 1994 Lakeview Dam Earthwork - RC-Lakeview-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Winchester Line F - RC-Winchester-1-6-1994 -- (RCFCWCD) 4 images
- 1995 Moreno Master Drainage Plan, Line 1-RC-1-7-1995 -- (RCFCWCD) 7 images
- 1995 Moreno Valley - WO-8-95-1995 -- (RCFCWCD) 4 images
- 1996 March Air Force Base Ortho - RC-1-72-1996 -- (RCFCWCD) 72 images
- 1996 Menifee Road - RC-Menifee-1-16-1996 -- (RCFCWCD) 16 images
- 1996 Moreno Main Drain Pipe Nason Retention Basin - RC-Moreno-1-6-1996 -- (RCFCWCD) 6 images
- 1996 Perris Valley Channel - RC-Perris-1-18-1996 -- (RCFCWCD) 19 images
- 1997 Bautista Basin - RC-1-7-1997 -- (RCFCWCD) 7 images
- 1997 Bautista Basin - RC-Bautista-1-13-1997 -- (RCFCWCD) 13 images
- 1997 Bradley Basin - RC-Bradley-1-2-1997 -- (RCFCWCD) 7 images
- 1997 Moreno F2 - RC-Moreno-1-3-1997 -- (RCFCWCD) 2 images
- 1997 Perris Lateral A - RC-1-15-1997 -- (RCFCWCD) 15 images
- 1997 Salt Creek Channel - RC-1-45-1997 -- (RCFCWCD) 45 images
- 1997 West Perris Ortho - RC-1-74-1997 -- (RCFCWCD) 72 images
- 1998 Bradley Road Channel - RC-1-17-1998 -- (RCFCWCD) 17 images
- 1998 Lakeview Dam - RC-Lakeview-1-10-1998 -- (RCFCWCD) 10 images
- 1998 Moreno Line F-2 - RC-Moreno-1-4-1998 -- (RCFCWCD) 4 images
- 1999 Bautista Basin / Inlet - RC-Bautista-1-14-1999 -- (RCFCWCD) 14 images
- 1999 Bradley Road Channel - RC-Bradley-1-15-1999 -- (RCFCWCD) 15 images
- 2000 Bradley Road Channel - RC-Bradley-1-16-2000 -- (RCFCWCD) 16 images
- 2000 Moreno Line F-2 - RC-Moreno-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Perris Valley Lateral A - RC-Perris-1-17-2000 -- (RCFCWCD) 17 images
- 2001 Bautista Basin - RC-1-11-2001 -- (RCFCWCD) 11 images
- 2001 Bradley Road Channel - RC-Bradley-1-15-2001 -- (RCFCWCD) 15 images
- 2001 Moreno Line F-2 - RC-Moreno-1-5-2001 -- (RCFCWCD) 5 images
- 2001 Moreno Valley South - RC-1-52-2001 -- (RCFCWCD) 53 images
- 2002 Little Lake Main Drain Pipe Line B - RC-Little-1-17-2002 -- (RCFCWCD) 17 images
- 2003 Bradley Basin - RC-Bradley2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Bradley Basin Annual - RC-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Buena Vista Basin - RC-Buena2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Buena Vista Basin - RC-Buena-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Canyon Springs Basin - RC-Canyon2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Canyon Springs Basin - RC-Canyon-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Lakeview Dam - RC-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Lasselle Basin - RC-Lassell-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Lasselle Basin - RC-Lasselle2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Little Lake Basin - RC-Little-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Little Lake Basin - RC-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Metz Basin - RC-Metz-1-6-2003 -- (RCFCWCD) 6 images
-
2003 Metz Basin - RC-Metz2-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Moreno Line F-2 - RC-Moreno-1-3-2003 -- (RCFCWCD) 2 images
- 2003 Nason Basin - RC-Nason2-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Nason Basin - RC-Nason-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Nason Basin - RC-Nason3-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Park Hill Basin - RC-Park-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Park Hill Basin - RC-Park2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Perris Basin - RC-Perris5-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Perris Basin - RC-Perris2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Perris Valley Channel Lateral A - RC-1-16-2003 -- (RCFCWCD) 16 images
- 2003 Perris Valley Channel Lateral B - RC-1-12-2003 -- (RCFCWCD) 12 images
- 2003 Perris Valley Lateral A - RC-Perris3-1-16-2003 -- (RCFCWCD) 16 images
- 2003 Perris Valley Lateral B - RC-Perris4-1-12-2003 -- (RCFCWCD) 12 images
- 2003 Perris Valley Storm Drain - RC-1-41-2003 -- (RCFCWCD) 41 images
- 2003 Perris Valley Storn Drain Channel - RC-1-42-2003 -- (RCFCWCD) 42 images
- 2003 Perris Third Street Basin - RC-Perris6-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Perris Third Street Basin - RC-Perris-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Pigeon Pass Dam - RC-1-15-2003 -- (RCFCWCD) 15 images
- 2003 Pigeon Pass Dam - RC-Pigeon2-1-15-2003 -- (RCFCWCD) 15 images
- 2003 Salt Creek Channel - RC-Salt-1-29-2003 -- (RCFCWCD) 29 images
- 2003 Salt Creek Reach 4 - RC-1-19-2003 -- (RCFCWCD) 19 images
- 2003 Salt Creek Reaches I - RC-1-18-2003 -- (RCFCWCD) 18 images
- 2003 Salt Creek Reaches II - RC-1-25-2003 -- (RCFCWCD) 25 images
- 2003 Salt Creek Reaches III - RC-1-31-2003 -- (RCFCWCD) 31 images
- 2003 San Jacinto Drain - RC-Jacinto-1-9-2003 -- (RCFCWCD) 9 images
- 2003 Towngate Basin - RC-Towngate2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Towngate Basin - RC-Towngate-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Winchester Ortho - RC-1-71-2003 -- (RCFCWCD) 70 images
- 2004 Bautista Basin - RC-Bautista-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Canyon Springs Basin - RC-Canyon-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Lasalle Basin - RC-Lasalle-1-2-2004-- (RCFCWCD) 2 images
- 2004 Menifee - Hawthorne Avenue Drain - RC-Menifee-1-13-2004 -- (RCFCWCD) 13 images
- 2004 Moreno Line F-2 - RC-Moreno-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Nason Basin - RC-Nason-1-4-2004 -- (RCFCWCD) 4 images
- 2004 Pigeon Pass Dam - RC-1-15A-2004 -- (RCFCWCD) 15 images
- 2004 Salt Creek Reach I - RC-1-18-2004 -- (RCFCWCD) 18 images
- 2004 Salt Creek Reach II - RC-1-25-2004-- (RCFCWCD) 25 images
- 2004 Salt Creek Reach III - RC-Salt-1-31-2004 -- (RCFCWCD) 31 images
- 2004 Sun City Ortho - RC-1-96-2004 -- (RCFCWCD) 96 images
- 2004 Towngate Basin - RC-Towngate-1-2-2004 -- (RCFCWCD) 2 images
- 2005 Badlands Ortho - RC-Badlands7-1-50-2005 -- (RCFCWCD) 50 images
- 2005 Bautista Basin - RC-Bautista7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 BP John Recycling - RC-BP-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Bradley Basin - RC-Bradley7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Bradley Basin - RC-Bradley-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Buena Vista Basin - RC-Buena7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Buena Vista Basin - RC-Buena-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Canyon Springs Basin - RC-Canyon7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Double Butte - RC-Double-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Lakeview Dam - RC-Lakeview7-1-4-2005 -- (RCFCWCD) 4 images
- 2005 Lasalle Basin - RC-Lasalle7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Little Lake Basin -RC-Little-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Little Lake Basin - RC-Little7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Menifee Hawthorne - RC-Menifee-1-7-2005 -- (RCFCWCD) 7 images
- 2005 Metz Basin - RC-Metz7-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Metz Basin - RC-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Moreno Line F-2 - RC-Moreno7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Moreno Main Drain Pipe Line K - RC-Moreno-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Nason Basin - RC-Nason-1-4-2005 -- (RCFCWCD) 4 images
- 2005 Nason Basin - RC-A-D-2005 -- (RCFCWCD) 3 images
- 2005 Noble Creek North - RC-Noble-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Noble Creek South - RC-Noble2-1-15-2005 -- (RCFCWCD) 15 images
- 2005 Park Hill Basin - RC-Park7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Park Hill Basin - RC-Park-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Perris Valley Storm Drain - RC-Perris7-1-25-2005 -- (RCFCWCD) 24 images
- 2005 Perris Basin - RC-Perris8-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Perris Basin - RC-Perris9-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Perris Valley Storm Drain Lateral A - RC-PerrisA-1-17-2005 -- (RCFCWCD) 17 images
- 2005 Perris Valley Storm Drain Lateral B - RC-PerrisB-1-12-2005 -- (RCFCWCD) 12 images
- 2005 Perris Basin - RC-Perris-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Perris Third Street Basin - RC-Perris2-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Perris Valley Storm Drain - RC-1-44-2005 -- (RCFCWCD) 44 images
- 2005 Perris Valley Street Drain Lateral A - RC-1-17-2005 -- (RCFCWCD) 17 images
- 2005 Pigeon Pass Dam - RC-Pigeon7-1-15-2005 -- (RCFCWCD) 15 images
- 2005 Towngate Basin - RC-Towngate7-1-2-2005 -- (RCFCWCD) 2 images
- 2006 Bautista Basin - RC-Bautista-1-3-2006 -- (RCFCWCD) 3 images
- 2006 BP John Recycling - RC-John-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Bradley Basin - RC-Bradley-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Buena Vista Basin - RC-Buena-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Canyon Springs Basin - RC-Canyon-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Double Butte Sanitary Landfill - RC-Spot-2006 -- (RCFCWCD) 1 image
- 2006 Lake Perris Ortho - RC-Perris-1-18-2006 -- (RCFCWCD) 18 images
- 2006 Lakeview Dam - RC-Lakeview-1-4-2006 -- (RCFCWCD) 4 images
- 2006 Lasalle Basin - RC-Lasalle-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Little Lake Basin - RC-Little2-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Metz Basin - RC-Metz6-1-6-2006 -- (RCFCWCD) 6 images
- 2006 Moreno Line F-2 - RC-Moreno-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Nason Basin - RC-Nason-1-4-2006 -- (RCFCWCD) 4 images
- 2006 Nuevo Channel - RC-Nuevo-1-7-2006 -- (RCFCWCD) 7 images
- 2006 Park Hill Basin - RC-Park-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Perris Valley Lateral B - RC-Perris9-1-12-2006 -- (RCFCWCD) 12 images
- 2006 Perris Valley Storm Drain - RC-1-41-2006 -- (RCFCWCD) 41 images
- 2006 Perris Valley Storm Drain Lateral A - RC-Perris7-1-16-2006 -- (RCFCWCD) 16 images
- 2006 Perris Third Street - RC-Perris8-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Pigeon Pass Dam - RC-Pigeon6-1-15-2006 -- (RCFCWCD) 15 images
- 2006 Pigeon Pass Ortho - RC-1-49-2006 -- (RCFCWCD) 49 images
- 2006 Salt Creek Reach 1 - RC-1-18-2006 -- (RCFCWCD) 18 images
- 2006 Salt Creek Reach I - RC-1-32-2006 -- (RCFCWCD) 32 images
- 2006 Salt Creek Reach II - RC-Salt6-1-25-2006 -- (RCFCWCD) 25 images
- 2006 Salt Creek Reach II - RC-1-25-2006 -- (RCFCWCD) 25 images
- 2006 Salt Creek Reach III - RC-1-31-2006 -- (RCFCWCD) 31 images
- 2006 Salt Creek Reach III -RC-Salt7-1-17-2006 -- (RCFCWCD) 17 images
- 2006 Towngate Basin - RC-Towngate-1-2-2006 -- (RCFCWCD) 2 images
- 2007 Bautista Basin - RC-Bautista-1-3-2007 -- (RCFCWCD) 2 images
- 2007 Bradley Basin - RC-Bradley-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Buena Vista Basin - RC-Buena-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Canyon Springs Basin - RC-Canyon-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Laselle Basin - RC-Laselle2-1-2-2007 -- (RCFCWCD) 2 images
- 2007 March Ortho - RC-1-62-2007 -- (RCFCWCD) 62 images
- 2007 Metz Basin - RC-Metz-1-5-2007 -- (RCFCWCD) 5 images
- 2007 Nason Basin - RC-Nason-1-4-2007 -- (RCFCWCD) 4 images
- 2007 Park Hill Basin - RC-Park-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Perris Basin -RC-Perris-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Perris Third Street Basin - RC-Perris2-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Pigeon Pass Dam - RC-1-15-2007 -- (RCFCWCD) 15 images
- 2007 Towngate Basin - RC-Towngate-1-2-2007 -- (RCFCWCD) 2 images
- 2008 Bautista Basin - RC-Bautista7-1-3-2008 -- (RCFCWCD) 3 images
- 2008 Buena Vista Basin - RC-Buena7-1-2-2008 -- (RCFCWCD) 2 images
-
2008 Canyon Springs Basin - RC-Canyon7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Lakeview Dam - RC-1-4-2008 -- (RCFCWCD) 4 images
- 2008 Lakeview Dam - RC-Lakeview7-1-4-2008 -- (RCFCWCD) 4 images
- 2008 Laselle Basin - RC-Laselle7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Metz Basin - RC-Metz7-1-6-2008 -- (RCFCWCD) 6 images
- 2008 Moreno Line F-2 - RC-Moreno7-1-3-2008 -- (RCFCWCD) 3 images
- 2008 Moreno Line F-2 - RC-1-3-2008 -- (RCFCWCD) 3 images
- 2008 Nason Basin - RC-Nason7-1-4-2008 -- (RCFCWCD) 4 images
- 2008 Palomar Detention Basin - RC-Palomar2-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Palomar Detention Basin - RC-Palomar-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Park Hill Basin - RC-Park7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Perris Basin - RC-Perris4-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Perris Third Street Basin - RC-Perris9-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Perris Valley Storm Drain - RC-Perris1-1-41-2008 -- (RCFCWCD) 41 images
- 2008 Perris Valley Storm Drain Lateral A -RC-Perris8-1-16-2008 -- (RCFCWCD) 15 images
- 2008 Perris Valley Storm Drain Lateral B - RC-Perris7-1-12-2008 -- (RCFCWCD) 13 images
- 2008 Perris Valley Storm Drain - RC-1-41-2008 -- (RCFCWCD) 41 images
- 2008 Perris Valley Storm Drain Lateral B - RC-1-12-2008 -- (RCFCWCD) 12 images
- 2008 Perris Valley Storm Drain Lateral A - RC-1-16-2008 -- (RCFCWCD) 16 images
- 2008 Pigeon Pass Dam - RC-Pigeon7-1-15-2008 -- (RCFCWCD) 15 images
- 2008 Salt Creek Reach I - RC-1-18-2008 -- (RCFCWCD) 18 images
- 2008 Salt Creek Reach II - RC-1-25-2008 -- (RCFCWCD) 25 images
- 2008 Salt Creek Reach III - RC-1-31-2008 -- (RCFCWCD) 30 images
- 2008 Towngate Basin - RC-Towngate7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Zone 4, Block 1 Ortho - RC-1-252-2008A -- (RCFCWCD) 251 images
- 2008 Zone 4, Block 1 Ortho - RC-253-255-2008B -- (RCFCWCD) 2 images
- 2009 Bradley Basin - RC-Bradley-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Bradley Basin - RC-Bradley7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Buena Vista Basin - RC-Buena7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Canyon Springs Basin - RC-Canyon7-1-2-2009-- (RCFCWCD) 2 images
- 2009 Lakeview Dam - RC-Lakeview7-1-4-2009 -- (RCFCWCD) 4 images
- 2009 Lasselle Basin - RC-Lasselle7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Metz Basin - RC-Metz7-1-6-2009 -- (RCFCWCD) 6 images
- 2009 Moreno Line F-2 - RC-Moreno7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Nason Basin - RC-Nason-1-4-2009 -- (RCFCWCD) 4 images
- 2009 Park Hill Basin - RC-Park7-1-2-2009 -- (RCFCWCD 2 images
- 2009 Perris Basin - RC-Perris8-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Perris Third Street Basin - RC-Perris7-1-2-2009 -- (RCFCWCD) 7 images
- 2009 Perris Valley Storm Drain - RC-1-41-2009 -- (RCFCWCD) 41 images
- 2009 Perris Valley Storm Drain Lateral A - RC-PerrisA-1-16-2009 -- (RCFCWCD) 16 images
- 2009 Perris Valley Storm Drain Lateral B - RC-PerrisB-1-12-2009 -- (RCFCWCD) 12 images
- 2009 Pigeon Pass Dam - RC-Pigeon7-1-15-2009 -- (RCFCWCD) 15 images
- 2009 Potrero Creek Basin - RC-Potrero-1-6-2009 -- (RCFCWCD) 6 images
- 2009 Salt Creek Channel - RC-Salt5-1-8-2009 -- (RCFCWCD) 8 images
- 2009 Salt Creek Reach I - RC-Salt9-1-18-2009 -- (RCFCWCD) 18 images
- 2009 Salt Creek Reach I - RC-1-18-2009 -- (RCFCWCD) 18 images
- 2009 Salt Creek Reach II - RC-1-25-2009 -- (RCFCWCD) 25 images
- 2009 Salt Creek Reach II - RC-Salt7-1-25-2009 -- (RCFCWCD) 25 images
- 2009 Salt Creek Rerach III - RC-Salt8-1-31-2009 -- (RCFCWCD) 31 images
- 2009 Salt Creek Reach III - RC-1-31-2009 -- (RCFCWCD) 31 images
- 2009 San Jacinto River - RC-1-34-2009 -- (RCFCWCD) 34 images
- 2009 Towngate Basin - RC-Towngate-1-2-2009 -- (RCFCWCD) 2 images
- 2010 Homeland - RC-1-6-2010 -- (RCFCWCD) 6 images
- 2010 Menifee - RC-1-30-2010 -- (RCFCWCD) 30 images
- 2010 Warm Springs Creek - RC-Warm-1-8-2010 -- (RCFCWCD) 8 images
- 2011 Bradley Basin - RC-Bradley-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Buena Vista Basin - RC-Buena-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Canyon Spring Basin - RC-Canyon-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Lakeview Dam - RC-Lakeview-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Lasalle Basin - RC-Lasalle-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Metz Basin - RC-Metz-1-6-2011 -- (RCFCWCD) 6 images
- 2011 Moreno Line F-2 - RC-MorenoF-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Moreno Main Drain Pipeline Sinclair Basin - RC-Moreno-1-10-2011 -- (RCFCWCD) 10 images
- 2011 Nason Basin - RC-Nason-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Palomar Detention Basin - RC-Palomar-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Park Hill Basin - RC-Park-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Perris Basin - RC-Perris3-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Perris Ortho - RC-1-197-2011 -- (RCFCWCD) 197 images
- 2011 Perris Third Street Basin - RC-Perris5-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Perris Valley Storm Drain - RC-20-41-2011 -- (RCFCWCD) 22 images
- 2011 Perris Valley Storm Drain - RC-Perris-4-1-19-2011 -- (RCFCWCD) 19 images
- 2011 Perris Valley Storm Drain Lateral A - RC-PerrisA-1-16-2011 -- (RCFCWCD) 2 images
- 2011 Perris Valley Storm Drain Lateral B - RC-1-12-2011 -- (RCFCWCD) 12 images
- 2011 Pigeon Pass Dam - RC-Pigeon-1-15-2011 -- (RCFCWCD) 15 images
- 2011 Potrero Creek Basin - RC-Potrero-1-6-2011 -- (RCFCWCD) 6 images
- 2011 Salt Creek Reach I - RC-Salt2-1-18-2011 -- (RCFCWCD) 18 images
- 2011 Salt Creek Reach III - RC-Salt3-1-31-2011 -- (RCFCWCD) 31 images
- 2011 Salt Creek Reach II - RC-SaltA-1-25-2011 -- (RCFCWCD) 25 images
- 2011 Sunnymead Main Drain Pipeline Line P6 - RC-Sunnymead-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Valle Vista Channel, Stage 11 - RC-Valle-1-6-2011 -- (RCFCWCD) 6 images
- 2012 Agriscape Lakeview - RC-Agriscape-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Agriscape Lakeview - RC-Agriscape7-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Bradley Basin - RC-Bradley7-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Buena Vista Basin - RC-Buena7-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Lakeview Dam - RC-Lakeview7-1-4-2012 -- (RCFCWCD) 4 images
- 2012 Lakeview Nuevo Main Drain Pipeline Lateral D - RC-LakeviewD-1-26-2012 -- (RCFCWCD) 26 images
- 2012 Lassalle Basin - RC-Lassalle7-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Metz Basin - RC-Metz7-1-6-2012 -- (RCFCWCD) 6 images
- 2012 Moreno Line F-2 - RC-Moreno7-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Nason Basin - RC-Nason7-1-4-2012 -- (RCFCWCD) 4 images
- 2012 Nason Basin - RC-Nason8-1-4-2012 -- (RCFCWCD) 4 images
- 2012 Palomar Detention Basin - RC-Palomar7-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Park Hill Basin - RC-Park7-1-2-2012 -- (RCFCWCD) 4 images
- 2012 Perris Basin - RC-Perris7-1-2-2012 -- (RCFCWCD) 4 images
- 2012 Perris Valley Storm Drain - RC-1-41-2012 -- (RCFCWCD) 41 images
- 2012 Perris Valley Storm Drain Lateral A - RC-1-16-2012 -- (RCFCWCD) 16 images
- 2012 Perris Valley Storm Drain Lateral B - RC-1-12-2012 -- (RCFCWCD) 12 images
- 2012 Pigeon Pass Dam - RC-Pigeon7-1-15-2012 -- (RCFCWCD) 15 images
- 2012 Potrero Creek Basin - RC-Potrero7-1-6-2012-- (RCFCWCD) 6 images
- 2012 Salt Creek Reach I - RC-Salt7-1-18-2012 -- (RCFCWCD) 18 images
- 2012 Salt Creek Reach II - RC-Salt8-1-25-2012 -- (RCFCWCD) 25 images
- 2012 Salt Creek Reach III - RC-1-31-2012 -- (RCFCWCD) 31 images
- 2013 Agriscape Lakeview - RC-Lakeview-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Bradley Basin - RC-Bradley-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Buena Vista Basin - RC-Buena-1-2-2013 -- (RCFCWCD) 2 image
- 2013 Canyon Springs Basin - RC-Canyon-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Indian Detention Basin - RC-Indian-1-4-2013 -- (RCFCWCD) 4 images
- 2013 Lasselle Basin - RC-Lasselle-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Little Lake Basin - RC-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Mead Valley Sanitary Landfill - RC-1-12-2013 -- (RCFCWCD) 12 images
- 2013 Mead Valley Sanitary Landfill Spot - RC-A-2013 -- (RCFCWCD) 1 images
- 2013 Metz Basin - RC-Metz-1-6-2013 -- (RCFCWCD) 6 images
- 2013 Moreno Line F-2 - RC-MorenoF-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Nason Basin -RC-Nason-1-4-2013 -- (RCFCWCD) 4 images
- 2013 Nuevo Channel - RC-Nuevo-1-6-2013 -- (RCFCWCD) 6 images
- 2013 Palomar Detention Basin - RC-Palomar-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Perris Basin - RC-Perris5-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Perris Third Street - RC-Perris3-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Perris Valley Storm Drain - RC-1-41-2013-- (RCFCWCD) 41 images
- 2013 Perris Valley Storm Drain Lateral A - RC-1-16-2013 -- (RCFCWCD) 16 images
- 2013 Perris Valley Storm Drain Lateral B - RC-PerrisB-1-12-2013 -- (RCFCWCD) 12 images
- 2013 Pigeon Pass Dam - RC-1-15-2013 -- (RCFCWCD) 15 images
- 2013 Potrero Creek Basin - RC-1-6-2013 -- (RCFCWCD) 6 images
- 2013 Salt Creek Reach I - RC-Salt-1-18-2013 -- (RCFCWCD) 18 images
- 2013 Salt Creek Reach II - RC-Salt2-1-25-2013-- (RCFCWCD) 25 images
- 2013 Salt Creek Reach III - RC-1-31-2013 -- (RCFCWCD) 31 images
- 2013 Towngate Basin - RC-Town-1-2-2013 -- (RCFCWCD) 2 images
- 2014 Lakeview / Nuevo Main Drain Pipeline Lateral D - RC-1-5-2014 -- (RCFCWCD) 5 images
- 2014 Lakeview Dam - RC-Lakeview-1-4-2014 -- (RCFCWCD) 4 images
- 2014 Metz Basin - RC-Metz-1-6-2014 -- (RCFCWCD) 6 images
- 2014 Nuevo Channel - RC-Nuevo-1-6-2014 -- (RCFCWCD) 6 images
- 2014 Perris Third Street Basin - RC-1-2-2014 -- (RCFCWCD) 2 images
- 2014 Perris Valley Storm Drain - RC-1-41-2014 -- (RCFCWCD) 41 images
- 2015 Bautista Basin - RC-Bautista-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Bradley Basin - RC-Bradley-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Buena Vista Basin - RC-Buena-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Canyon Springs Basin - RC-Canyon-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Indian Detention Basin - RC-Indian2-1-4-2015 -- (RCFCWCD) 4 images
- 2015 Lakeview Dam - RC-Lakeview-1-4-2015 -- (RCFCWCD) 4 images
- 2015 Lasalle Basin - RC-Lasalle-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Little Lake Basin - RC-Little2-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Metz Basin - RC-Metz-1-6-2015 -- (RCFCWCD) 6 images
- 2015 Moreno Line F-2 - RC-MorenoF-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Nason Basin - RC-Nason-1-4-2015 -- (RCFCWCD) 4 images
- 2015 Nuevo Channel - RC-Nuevo-1-6-2015 -- (RCFCWCD) 6 images
- 2015 Palomar Basin - RC-Palomar-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Park Hill Basin - RC-Park-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Perris Basin - RC-Perris2-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Perris Third Street Basin - RC-Perris-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Perris Lateral A- RC-1-16-2015 -- (RCFCWCD) 16 images
- 2015 Perris Lateral B - RC-1-12-2015 -- (RCFCWCD) 12 images
- 2015 Perris Valley Storm Drain - RC-1-41-2015 -- (RCFCWCD) 41 images
- 2015 Pigeon Pass Dam - RC-Pigeon-1-15-2015 -- (RCFCWCD) 15 images
- 2015 Portrero Creek Basin - RC-Portrero-1-6-2015 -- (RCFCWCD) 6 images
- 2015 Salt Creek / Evans Road Basin - RC-Evans-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Salt Creek Reach I - RC-1-18-2015 -- (RCFCWCD) 18 images
- 2015 Salt Creek Reach II - RC-Salt-1-25-2015 -- (RCFCWCD) 25 images
- 2015 Salt Creek Reach III - RC-1-31-2015 -- (RCFCWCD) 31 images
- 2015 Towngate Basin - RC-Towngate-1-2-2015 -- (RCFCWCD) 2 images
-
Palm Springs (11,269 images)
Cities in the eastern portion of Riverside County called the Coachella Valley include Desert Hot Springs, Palm Springs, Rancho Mirage, Cathedral City, Palm Desert, La Quinta, Indian Wells, Coachella, Indio, Mecca, Oasis, and Thermal. The northern portion of the Salton Sea is also included.
- 1948 Cathedral Canyon - RC-1-27-1948 -- (RCFCWCD) 21 images
- 1948 Cathedral City - RC-18-26-1948 -- (RCFCWCD) 9 images
- 1948 Cathedral City Obliques - RC-8-17-1948 -- (RCFCWCD) 10 images
- 1948 Desert Hot Springs - RC-5-27-1948 -- (RCFCWCD) 5 images
- 1948 Palm Springs - RC-1-71-1948 -- (RCFCWCD) 66 images
- 1948 Palm Springs - RC-1-22-1948 -- (RCFCWCD) 18 images
- 1948 Palm Springs & Chino Cove - RC-79-111-1948 -- (RCFCWCD) 24 images
- 1948 Whitewater River - RC-1-5-1948 -- (RCFCWCD) 5 images
- 1948 Whitewater River at Cathedral City - RC-1-7-1948 -- (RCFCWCD) 7 images
- 1949 Long Canyon, Wide Canyon, Mission Creek, Whitewater River - RC-40-72-1949 -- (RCFCWCD) 17 images
- 1949 Palm Canyon Book 13A - RC-AXM-F-20F-109-113-1949 -- (RCFCWCD) 7 images
- 1949 Palm Canyon Book 13A - RC-AXM-F-21F-26-28-1949 -- (RCFCWCD) 7 images
- 1949 Palm Springs Book 14A - RC-AXM-F-15F-62-219-1949 -- (RCFCWCD) 36 images
- 1949 Whitewater River Obliques - RC-1-7-1949 -- (RCFCWCD) 7 images
- 1953 Desert Hot Springs Book 8B - RC-AXM-1K-24-63-1953 -- (RCFCWCD) 18 images
- 1953 Desert Hot Springs Book 4B - RC-AXM-3K-3-15-1953 -- (RCFCWCD) 7 images
- 1953 Desert Hot Springs Book 9B -RC-AXM-9K-53-73-1953 -- (RCFCWCD) 11 images
- 1953 Desert Hot Springs Book 10K-5 - RC-AXM-10K-5-25-1953 -- (RCFCWCD) 12 images
- 1953 Desert Hot Springs Book 8B -RC-AXM-13K-156-188-1953 -- (RCFCWCD) 14 images
- 1954 Desert Hot Springs Book 9B - RC-AXM-15K-179-181-1954 -- (RCFCWCD) 2 images
- 1955 Cathedral City/ Palm Springs - RC-1-1-3-4 -- (RCFCWCD) 6 images
- 1955 Desert Hot Springs - RC-4-1-9-6 -- (RCFCWCD) 45 images
- 1957 Desert Hot Springs - RC-4-1-68 -- (RCFCWCD) 68 images
- 1958 East & West Wide Canyon - RC-31-50 -- (RCFCWCD) 21 images
- 1958 Long Canyon - RC-52-57 -- (RCFCWCD) 6 images
- 1959 Mission Creek - RC-23-41-1959 -- (RCFCWCD) 19 images
- 1959 Mission Creek Watershed - RC-23-49 -- (RCFCWCD) 8 images
- 1959 Mission Creek Watershed - RC-80-109 -- (RCFCWCD) 30 images
- 1959 Mission Creek Watershed - RC-110-138 -- (RCFCWCD) 29 images
- 1959 Mission Creek Watershed - RC-143-158 -- (RCFCWCD) 16 images
- 1959 Mission Creek - RC-50-79 -- (RCFCWCD) 30 images
- 1959 Mission Creek & Morongo Canyon - RC-1-44 -- (RCFCWCD) 27 images
- 1959 Palm Springs - Chino Canyon-RC-33-72 -- (RCFCWCD) 40 images
- 1959 Salton Sea - RC-1-82 -- (RCFCWCD) 82 images
- 1959 Tahquitz Creek - RC-1-17 -- (RCFCWCD) 82 images
- 1959 Willow Hole/7 Palms Valley - RC-1-22 -- (RCFCWCD) 20 images
- 1960 Blythe Airport - RC-1-8 -- (RCFCWCD) 8 images
- 1960 Desert Hot Springs - RC-12 -- (RCFCWCD) 1 image
- 1960 Mission Creek - RC-25-27 -- (RCFCWCD) 3 images
- 1960 Palm Springs - RC-13-16 -- (RCFCWCD) 4 images
- 1960 Thermal Airport - RC-9-11 -- (RCFCWCD) 3 images
- 1960 Whitewater - RC-27-46 -- (RCFCWCD) 20 images
- 1960 Whitewater - RC-31-57 -- (RCFCWCD) 27 images
- 1961 Cathedral City - RC-23-25-1961 -- (RCFCWCD) 3 images
- 1962 Fun Valley - RC-34-64-1962 -- (RCFCWCD) 31 images
- 1963 Desert Hot Springs - RC-1-41-1963 -- (RCFCWCD) 40 images
- 1964 Palm & Chino Canyon - RC-23-56-1964 -- (RCFCWCD) 29 images
- 1965 Chino Canyon - RC-70-77-1965 -- (RCFCWCD) 8 images
- 1965 Desert Hot Springs Convergents - RC-1-5-1965 -- (RCFCWCD) 4 images
- 1965 Mission Creek - TC-78-158-1965 -- (RCFCWCD) 80 images
- 1965 Palm Canyon Wash- RC-50-69-1965 -- (RCFCWCD) 20 images
- 1965 Palm Canyon - RC-121-143-1965 -- (RCFCWCD) 24 images
- 1965 Palm Springs - RC-1-119-1965 -- (RCFCWCD) 122 images
- 1965 Tahquitz Creek - RC-1-49-1965 -- (RCFCWCD) 46 images
- 1966 Palm Canyon - RC-21-43-1966 -- (RCFCWCD) 23 images
- 1966 Palm Canyon - RC-31-60-1966 -- (RCFCWCD) 30 images
- 1967 Cathedral City - RC-A-6-17-1967 -- (RCFCWCD) 13 images
- 1967 Dillon Road - RC-11-18-1967 -- (RCFCWCD) 8 images
- 1967 Palm Canyon - RC-1-9-1967 -- (RCFCWCD) 9 images
- 1967 San Gorgonio River - Cabazon - RC-1-1-1-7-1967 -- (RCFCWCD) 7 images
- 1968 Cathedral City - RC-32-41-1968 -- (RCFCWCD) 10 images
- 1968 Eagle Mountain Dump - RC-A-1968 -- (RCFCWCD) 1 image
- 1968 Indian Wells - RC-1-4-1968 -- (RCFCWCD) 4 images
- 1968 Palm Springs - RC-19-31-1968 -- (RCFCWCD) 13 images
- 1968 Wide Canyon - RC-12-18-1968 -- (RCFCWCD) 7 images
- 1969 Fann Hill - RC-20-51-1969 -- (RCFCWCD) 30 images
- 1969 Indian Wells -RC-3-5-1969 -- (RCFCWCD) 3 images
- 1969 Indio - RC-27-41-1969 -- (RCFCWCD) 15 images
- 1969 La Quinta - RC-1-2-1969 -- (RCFCWCD) 2 images
- 1969 Palm Canyon Dunn Road - RCPalm-1-8-1969 -- (RCFCWCD) 8 images
- 1969 Palm Springs - De Valle Drive - RC-19-1969 -- (RCFCWCD) 1 images
- 1969 Whitewater - RC-1-6-1969 -- (RCFCWCD) 6 images
- 1969 Whitewater -RC-1-11-1969 -- (RCFCWCD) 11 images
- 1970 Desert Hot Springs -RC-1-28-1970 -- (RCFCWCD) 28 images
- 1971 Bob Hope & Country Club Road - RC-13-1971 -- (RCFCWCD) 1 image
- 1971 Little Morongo - RC-1-45-1971 -- (RCFCWCD) 45 images
- 1971 Mission Creek - RC-18-71-1971 -- (RCFCWCD) 28 images
- 1971 Palm Springs Bypass/U.S. Hwy. 111 - RC-1-124-1971 -- (RCFCWCD) 124 images
- 1971 Pinyon Flat - RC-Pinyon-1-3-1971 -- (RCFCWCD) 3 images
- 1971 Tahquitz - RC-Tahquitz-1-11-1971 -- (RCFCWCD) 11 images
- 1971 Whitewater - RC-Whitewater-1-3-1971 -- (RCFCWCD) 3 images
- 1971 Whitewater River - RC-1-46-1971 -- (RCFCWCD) 46 images
- 1972 Coachella Fill - RC-5-13-1972 -- (RCFCWCD) 9 images
- 1972 Mission Creek - RC-20-55-1972 -- (RCFCWCD) 36 images
- 1972 Oasis Disposal - RC-3-7-1972 -- (RCFCWCD) 5 images
- 1972 Whitewater - RC-1-10-1972 -- (RCFCWCD) 9 images
- 1972 Windy Point - RC-23-39-1972 -- (RCFCWCD) 17 images
- 1973 Blythe Disposal - RC-Blythe-1-11-1973 -- (RCFCWCD) 11 images
- 1973 Edon Hill Fill - RC-80-84-1973 -- (RCFCWCD) 5 images
- 1973 Mecca Disposal - RC-1-4-1973 -- (RCFCWCD) 4 images
- 1973 Palm Springs Reservoir - RC-1-11-1973 -- (RCFCWCD) 11 images
- 1974 Cahuilla Hills Convergent - RC-1-18-1974 -- (RCFCWCD) 18 images
- 1974 Indio West Half 755R7E - RC-1-42-1975 -- (RCFCWCD) 81 images
- 1974 Indio Test - RC-1-5-1974 -- (RCFCWCD) 4 images
- 1974 Indio Test - RC-10-19-1974 -- (RCFCWCD) 10 images
- 1974 Indio Test - RC-28-39-1974 -- (RCFCWCD) 12 images
- 1974 Palm Administration Center - RC-Palm-Springs-1-4-1974 -- (RCFCWCD) 4
- 1974 Wide Canyon Channel -RC-1-79-1974 -- (RCFCWCD) 79 images
- 1975 Chino Ramp 5 - RC-1-23-1975 -- (RCFCWCD) 23 images
- 1975 Coachella - RC-1-5-1975 -- (RCFCWCD) 5 images
- 1975 Eagle Mountain - RC-1-4-1975 -- (RCFCWCD) 4 images
- 1975 Edom Hill - RC-Edom-1-4-1975 -- (RCFCWCD) 4 images
- 1975 Indio - RC-17-49-1975 -- (RCFCWCD) 12 images
- 1975 Indio - RC-50-61-1975 -- (RCFCWCD) 30 images
- 1975 La Quinta Ramp 3 - RC-28-74-1975 -- (RCFCWCD) 36 images
- 1975 Palm Springs Revision - RC-1-18-1975 -- (RCFCWCD) 17 images
- 1976 Blythe Disposal - RC-Blythe-3-5-1976 -- (RCFCWCD) 3 images
- 1976 Cathedral City - RC-1-1976 -- (RCFCWCD) 1 image
- 1976 Mecca Disposal - RC-Mecca-1-5-1976 -- (RCFCWCD) 5 images
- 1976 Oasis Disposal Convergents - RC-Oasis-1-5-1976 -- (RCFCWCD) 5 images
- 1976 Thousand Palms - RC-59-107-1976 -- (RCFCWCD) 48 images
- 1976 Thousand Palms Ortho - RC-108-125-1976 -- (RCFCWCD) 18 images
- 1976 Wide Canyon Dam - RC-1-8-1976 -- (RCFCWCD) 8 images
- 1977 Blythe Disposal - RC-3-5-1977 -- (RCFCWCD) 3 images
- 1977 Blythe Ramp 8 - RC-Blythe-1-23-1977 -- (RCFCWCD) 23 images
- 1977 Coachella Disposal - RC-Coachella-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Desert Center Disposal - RC-1-3-1977 -- (RCFCWCD) 3 images
- 1977 East Deception Canyon - RC-1-817-1977 - Part 1 -- (RCFCWCD) 252 images
- 1977 East Deception Canyon - RC-1-817-1977 - Part 2 -- (RCFCWCD) 252 images
- 1977 East Deception Canyon - RC-1-817-1977 - Part 3 -- (RCFCWCD) 252 images
- 1977 East Deception Canyon - RC-1-817-1977 - Part 4 -- (RCFCWCD) 59 images
- 1977 East Deception Canyon Low Obliques - RC-818-826-1977 -- (RCFCWCD) 9 images
- 1977 Edom Hill Disposal - RC-Edom-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Horace Miller Park - RC-Horace-3-5-1977 -- (RCFCWCD) 3 images
- 1977 Mayflower Park - RC-Mayflower-3-5-1977 -- (RCFCWCD) 2 images
- 1977 Mecca Disposal - RC-Mecca-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Oasis Disposal - RC-Oasis-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Palm Canyon Levee - RC-12-43-1977 -- (RCFCWCD) 32 images
- 1977 Palm Canyon Levee Ortho - RC-8-11-1977 -- (RCFCWCD) 4 images
- 1977 Palm Springs - Flight 7799 -- (Imbiorski) 37 images
- 1977 Peter McIntyre Park - RC-Peter-3-5-1977 -- (RCFCWCD) 3 images
- 1977 Thousand Palms - RC-1-01-0-02-1977 -- (RCFCWCD) 2 images
- 1977 Thousand Palms Flood Damage - RC-1-92-1977 -- (RCFCWCD) 92 images
- 1977 Wide Canyon - RC-Wide-1-13-1977 -- (RCFCWCD) 13 images
- 1977 Wide Canyon Dam Low Obliques - RC-14-17-1977 -- (RCFCWCD) 4 images
- 1977 Wonder Valley - RC-1-41-1977 -- (RCFCWCD) 41 images
- 1978 Blythe Disposal - RC-Blythe-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Coachella Disposal - RC-Coachella-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Desert Hot Springs - RC-1-10-1978 -- (RCFCWCD) 10 images
- 1978 Desert Hot Springs - RC-1-46-1978 -- (RCFCWCD) 46 images
- 1978 Edom Hill Disposal - RC-Edom-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Mecca Disposal - RC-Mecca-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Mission Creek - RC-1-36-1978 -- (RCFCWCD) 36 images
- 1978 Oasis Disposal - RC-Oasis-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Palm Canyon - RC-1-12-1978 -- (RCFCWCD) 12 images
- 1979 Badlands Sanitary Landfill - RC-4-7-12-1979 -- (RCFCWCD) 12 images
- 1979 Blythe - RC-2-3-5-1979 -- (RCFCWCD) 3 images
- 1979 Coachella Sanitary Landfill - RC-Coachella-1-1-7-1979 -- (RCFCWCD) 7 images
- 1979 Desert Hot Springs - RC-1-11-1979 -- (RCFCWCD) 11 images
- 1979 Desert Hot Springs - RC-Desert-1-11-1979 -- (RCFCWCD) 11 images
- 1979 Desert Hot Springs - RC-1-56-1979 -- (RCFCWCD) 57 images
- 1979 Mecca #1 Sanitary Landfill - RC-Mecca-3-5-7-1979 -- (RCFCWCD) 3 images
- 1979 Mecca #2 Sanitary Landfill - RC-Mecca-3-5-9-1979 -- (RCFCWCD) 5 images
- 1979 Palm Canyon - RC-1-6-1979 -- (RCFCWCD) 6 images
- 1979 Tahquitz Basin - RC-Tahquitz-5-7-1979 -- (RCFCWCD) 3 images
- 1979 Tahquitz Basin - RC-TahquitzBasin-1-5-1979 -- (RCFCWCD) 4 images
- 1979 Tahquitz Basin Obliques - RC-8-10-1979 -- (RCFCWCD) 3 images
- 1979 Tahquitz Creek - RC-4-7-15-1979 -- (RCFCWCD) 9 images
- 1979 Tahquitz Creek - RC-16-56-1979 -- (RCFCWCD) 40 images
- 1980 Desert Hot Springs - RC-1-94-1980 -- (RCFCWCD) 92 images
- 1980 Edom Hill - RC-5-14-1980 -- (RCFCWCD) 10 images
- 1980 Mission Creek - RC-Mission-1-35-1980 -- (RCFCWCD) 35 images
- 1980 Mission Creek & Little Morongo - RC-1-78-1980 -- (RCFCWCD) 78 images
- 1980 Noble Creek - RC-1-9-1980 -- (RCFCWCD) 9 images
- 1980 Palm Canyon - RC-Palm-Canyon-1-34-1980 -- (RCFCWCD) 34 images
- 1980 Palm Canyon Flood Damage - RC-1-34-1980 -- (RCFCWCD) 26 images
- 1980 Palm Canyon Wash - RC-1-38-1980 -- (RCFCWCD) 38 images
- 1980 Tahquitz Creek - RC-1-14-1980 -- (RCFCWCD) 14 images
- 1982 Desert Hot Springs -RC-1-48-1982 -- (RCFCWCD) 48 images
- 1982 Desert Hot Springs Ortho - RC-1-18-1982 -- (RCFCWCD) 18 images
- 1982 Minto Way - RC-12-19-1982 -- (RCFCWCD) 8 images
- 1982 Minto Way - RC-Minto-1-11-1982 -- (RCFCWCD) 11 images
- 1982 Tachevah Dam - RC-5-15-1982 -- (RCFCWCD) 11 images
- 1982 Verbena Storm Channel - RC-9-27-1982 -- (RCFCWCD) 19 images
- 1983 Blythe Sanitary Landfill - RC-3-5-1983 -- (RCFCWCD) 3 images
- 1983 Blythe Sanitary Landfill - RC-4-11-1983 -- (RCFCWCD) 17 images
- 1983 Camino Real Road - RC-Camino-1-32-1983 -- (RCFCWCD) 32 images
- 1983 Coachella - RC-5-12-1983 -- (RCFCWCD) 8 images
- 1983 Coachella - RC-Coachella-1-10-1983 -- (RCFCWCD) 8 images
- 1983 Colorado River - RC-17-1-21-1983 -- (RCFCWCD) 21 images
- 1983 Colorado River - RC-18-1-17-1983 -- (RCFCWCD) 17 images
- 1983 Colorado River - RC-19-1-23-1983 -- (RCFCWCD) 23 images
- 1983 Desert Center - RC-Desert-2-4-1983 -- (RCFCWCD) 3 images
- 1983 Desert Center - RC-Desert-3-5-1983 -- (RCFCWCD) 3 images
- 1983 Desert Center - RC-2-4-1983 -- (RCFCWCD) 3 images
- 1983 Desert Center Sanitary Landfill - RC-3-4-1983 -- (RCFCWCD) 2 images
- 1983 Edom Hill Sanitary Landfill - RC-1-14-1983 -- (RCFCWCD) 14 images
- 1983 Edom Hill Sanitary Landfill - RC-Edom-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Gentian Avenue Storm Drain - RC-1-10-1983 -- (RCFCWCD) 10 images
- 1983 Mecca II - RC-Mecca2-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Mecca II - RC-MeccaII-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Mecca II Disposal - RC-Mecca-3-6-1983 -- (RCFCWCD) 4 images
- 1983 Mecca Sanitary Landfill - RC-2-5-1983 -- (RCFCWCD) 3 images
- 1983 Morongo Channel - RC-1-20-1983 -- (RCFCWCD) 20 images
- 1983 Oasis Sanitary Landfill - RC-Oasis-1-3-1983 -- (RCFCWCD) 3 images
- 1983 Oasis Sanitary Landfill - RC-Oasis2-2-4-1983 -- (RCFCWCD) 3 images
- 1983 Oasis Sanitary Landfill - RC-Oasis-2-4-1983 -- (RCFCWCD) 3 images
- 1983 Palm Canyon - RC-1-13-1983 -- (RCFCWCD) 13 images
- 1983 Palm Canyon - RC-14-25-1983 -- (RCFCWCD) 12 images
- 1983 Tahchevah Sanitary Landfill - RC-Tahchevah-1-6-1983 -- (RCFCWCD) 6 images
- 1983 Verbana Extension - RC-3-8-1983 -- (RCFCWCD) 6 images
- 1983 Wide Canyon - RC-1-7-1983 -- (RCFCWCD) 7 images
- 1984 Blythe Sanitary Landfill - RC-Blythe-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Desert Center -RC-1-3-1984 -- (RCFCWCD) 3 images
- 1984 Desert Center Sanitary Landfill - RC-Desert-1-4-1984 -- (RCFCWCD) 3 images
- 1984 Desert Center Sanitary Landfill - RC-Desert2-1-3-1984 -- (RCFCWCD) 3 images
- 1984 East Cathedral Canyon - RC-Cathedral-1-11-1984 -- (RCFCWCD) 11 images
- 1984 Eagle Canyon - RC-1-10-1984 -- (RCFCWCD) 10 images
- 1984 Eagle Canyon Channel - RC-1-26-1984 -- (RCFCWCD) 26 images
- 1984 Mecca I Sanitary Landfill - RC-1-5-1984 -- (RCFCWCD) 5 images
- 1984 Mecca II Sanitary Landfill - RC-Mecca-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Mecca II Sanitary Landfill - RC-Mecca-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Mission Creek - RC-Mission-1-12-1984 -- (RCFCWCD) 12 images
- 1984 Oasis Sanitary Landfill - RC-Oasis-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Oasis Sanitary Landfill - RC-Oasis-1-5-1984 -- (RCFCWCD) 5 images
- 1984 Oasis Sanitary Landfill - RC-Oasis2-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Oasis Sanitary Landfill - RC-Oasis3-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Palm Canyon - RC-Palm-1-12-1984 -- (RCFCWCD) 12 images
- 1984 Palm Springs Lines 15, 15B, 15BA -RC-PS-1-26-1984 -- (RCFCWCD) 26 images
- 1984 Sunrise Way Storm Drain - RC-1-14-1984 -- (RCFCWCD) 14 images
- 1984 Sunrise Way Storm Drain - RC-1-46-1984 -- (RCFCWCD) 46 images
- 1984 Tachevah Basin - RC-Tahchevah-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Wide Canyon Basin - RC-Wide-1-6-1984 -- (RCFCWCD) 6 images
- 1985 Big Morongo Mapping Phase 1 - RC-1-19-1985 -- (RCFCWCD) 19 images
- 1985 Big Morongo Mapping Phase 1 - RC-1-31-1985 -- (RCFCWCD) 31 images
- 1985 Big Morongo Mapping Phase 2 - RC-1-17-1985 -- (RCFCWCD) 17 images
- 1985 Big Morongo Mapping Phase 3 - RC-51-529-1985 -- (RCFCWCD) 35 images
- 1985 Big Morongo Ortho Phase 1 - RC-1-14-1985 -- (RCFCWCD) 14 images
- 1985 Big Morongo Ortho Phase 2 - RC-Morongo-1-14-1985 -- (RCFCWCD) 14 images
- 1985 Chino Canyon - RC-1-11-1985 -- (RCFCWCD) 11 images
- 1985 Eagle Mountain Sanitary Landfill - RC-Eagle-1-3-1985 -- (RCFCWCD) 3 images
- 1985 East Cathedral Canyon - RC-East-1-17-1985 -- (RCFCWCD) 17 images
- 1985 Mecca 2 Sanitary Landfill - RC-Mecca2-1-4-1985 -- (RCFCWCD) 4 images
-
1985 Mission Creek - RC-1-38-1985 -- (RCFCWCD) 38 images
- 1985 Oak Street Basin - RC-Oak-1-4-1985 -- (RCFCWCD) 4 images
- 1985 Oasis Sanitary Landfill - RC-Oasis-1-3-1985 -- (RCFCWCD) 3 images
- 1985 Palm Canyon - RC-1-30-1985 -- (RCFCWCD) 30 images
- 1985 Sunrise Way Storm Drain - RC-1-2-1985 -- (RCFCWCD) 2 images
- 1985 Sunrise Way Storm Drain - RC-1-6-1985 -- (RCFCWCD) 6 images
- 1985 Tahchevah Basin - RC-Tahchevah-1-6-1985 -- (RCFCWCD) 6 images
- 1985 West Cathedral Canyon - RC-1-18-1985 -- (RCFCWCD) 18 images
- 1985 Wide Canyon Basin - RC-Wide-1-7-1985 -- (RCFCWCD) 7 images
- 1986 Blythe Disposal - RC-Blythe-1-3-1986 -- (RCFCWCD) 3 images
- 1986 Camino Real Storm Drain - RC-1-47-1986 -- (RCFCWCD) 46 images
- 1986 Camino Real Storm Drain Extension - RC-1-48-1986 -- (RCFCWCD) 48 images
- 1986 Coachella Valley Sanitary Landfill - RC-1-7-1986 -- (RCFCWCD) 7 images
- 1986 Big Morongo Ortho - RC-Morongo-1-3-1986 -- (RCFCWCD) 3 images
- 1986 Desert Center - RC-1-3-7-1986 -- (RCFCWCD) 4 images
- 1986 Eagle Canyon Storm Drain - RC-1-3-1986 -- (RCFCWCD) 3 images
- 1986 Mecca II Sanitary Landfill - RC-Mecca-1-7-1986 -- (RCFCWCD) 7 images
- 1986 Oasis - RC-Oasis-1-4-1986 -- (RCFCWCD) 4 images
- 1986 Palm Canyon - RC-1-28-1986 -- (RCFCWCD) 27 images
- 1986 Palm Valley Channel - RC-Palm-1-6-1986 -- (RCFCWCD) 6 images
- 1987 Blythe - RC-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Chino Canyon Leveee - RC-Chino-1-10-1987 -- (RCFCWCD) 10 images
- 1987 East & West Cathedral Canyon - RC-1-38-1987 -- (RCFCWCD) 38 images
- 1987 Mission Creek - RC-Mission-1-38-1987 -- (RCFCWCD) 38 images
- 1987 Palm Canyon - RC-1-30-1987 -- (RCFCWCD) 30 images
- 1987 Tachevah Basin - RC-Tahchevah-1-6-1987 -- (RCFCWCD) 6 images
- 1987 Whitewater River - RC-1-25-1987 -- (RCFCWCD) 25 images
- 1987 Whitewater River - RC-1-46-1987 -- (RCFCWCD) 46 images
- 1987 Whitewater River - RC-2-22-1987 -- (RCFCWCD) 10 images
- 1987 Whitewater River - RC-47-57-1987 -- (RCFCWCD) 11 images
- 1987 Whitewater Levee - RC-1-39-1987 -- (RCFCWCD) 39 images
- 1987 Wide Canyon Basin - RC-Wide-1-7-1987 -- (RCFCWCD) 6 images
- 1988 Cathedral Canyon - RC-1-32-1988 -- (RCFCWCD) 32 images
- 1988 Coachella Spot - RC-Coachella-1-1988 -- (RCFCWCD) 1 image
- 1988 Coachella St. - RC-Coachella2-1-9-1988 -- (RCFCWCD) 9 images
- 1988 Edom Hill Spot - RC-Edom-1-1988 -- (RCFCWCD) 1 image
- 1988 Edom Hill Sanitary Landfill - RC-Edom2-1-10-1988 -- (RCFCWCD) 10 images
- 1988 Farrell Basin - RC-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Mission Creek - RC-1-38-1988 -- (RCFCWCD) 38 images
- 1988 Palm Canyon - RC-1-29-1988 -- (RCFCWCD) 29 images
- 1988 Palm Springs Line 4 - RC-Palm-1-31-1988 -- (RCFCWCD) 31 images
- 1988 Tahchevah Basin - RC-Tahchevah-1-6-1988 -- (RCFCWCD) 6 images
- 1988 Tahquitz Creek - RC-11-18-1988 -- (RCFCWCD) 6 images
- 1988 Wide Canyon - RC-1-7-1988 -- (RCFCWCD) 7 images
- 1989 Blythe Sanitary Landfill - RC-Blythe-1-1989 -- (RCFCWCD) 1 image
- 1989 Blythe Sanitary Landfill - RC-Blythe2-1-3-1989 -- (RCFCWCD) 3 images
- 1989 Coachella Sanitary Landfill - RC-1-10-1989 -- (RCFCWCD) 10 images
- 1989 Coachella Sanitary Landfill Spot - RC-Coachella-1-1989 -- (RCFCWCD) 1 image
- 1989 Desert Center Spot - RC-1-1989 -- (RCFCWCD) 1 image
- 1989 Desert Center - RC-Desert-1-3-1989 -- (RCFCWCD) 3 images
- 1989 East & West Cathedra Canyon - RC-1-34-1989 -- (RCFCWCD) 34 images
- 1989 Edom Hill - RC-Edom2-1-10-1989 -- (RCFCWCD) 10 images
- 1989 Edom Hill Spot - RC-Edom-1-1989 -- (RCFCWCD) 1 image
- 1989 Mecca II Sanitary Landfill - RC-1-5-1989 -- (RCFCWCD) 5 images
- 1989 Mission Creek - RC-1-34-1989 -- (RCFCWCD) 34 images
- 1989 Morongo Wash - RC-1-28-1989 -- (RCFCWCD) 28 images
- 1989 Oasis Sanitary Landfill - RC-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Palm Canyon - RC-1-29-1989 -- (RCFCWCD) 29 images
- 1989 Tachevah Basin - RC-Tahchevah-1-6-1989 -- (RCFCWCD) 6 images
- 1989 Wide Canyon Basin - RC-Wide-1-6-1989 -- (RCFCWCD) 6 images
- 1990 Blythe - RC-1-3-1990 -- (RCFCWCD) 3 images
- 1990 Blythe Sanitary Landfill Spot - RC-1-1990 -- (RCFCWCD) 1 image
- 1990 Desert Center - RC-1-4-1990 -- (RCFCWCD) 4 images
- 1990 East & West Cathedral Canyon Channel - RC-1-36-1990 -- (RCFCWCD) 36 images
- 1990 Mecca - RC-1-5-1990 -- (RCFCWCD) 5 images
- 1990 Palm Canyon Channel - RC-1-30-1990 -- (RCFCWCD) 30 images
- 1990 Palm Canyon Extension - RC-1-7-1990 -- (RCFCWCD) 7 images
- 1990 Palm Springs Line 6 & 6B - RC-1-27-1990 -- (RCFCWCD) 27 images
- 1990 Palm Springs Line 34 - RC-1-24-1990 -- (RCFCWCD) 24 images
- 1990 Pinyon Flats - RC-Pinyon2-1-4-1990 -- (RCFCWCD) 4 images
- 1990 Pinyon Flats - RC-Pinion-1-3-1990 -- (RCFCWCD) 3 images
- 1990 Riverside / San Bernardino - RC-K33_204-K-1990 -- (RCFCWCD) 2 images
- 1990 Riverside / San Bernardino - RC-K33_204-O-1990 -- (RCFCWCD) 1 image
- 1990 Riverside / San Bernardino - RC-K33_204-P-1990 -- (RCFCWCD) 3 images
- 1990 Riverside / San Bernardino - RC-K33_204-Q-1990 -- (RCFCWCD) 3 images
- 1990 Riverside / San Bernardino - RC-K33_204-R-1990 -- (RCFCWCD) 3 images
- 1990 Riverside / San Bernardino - RC-K33_204-S-1990 -- (RCFCWCD) 3 images
- 1990 Stewart Park - RC-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Tahquitz Basin - RC-Tahquitz-1-1990 -- (RCFCWCD) 1 image
- 1990 Tahchevah Basin - RC-1-6-1990 -- (RCFCWCD) 6 images
- 1991 Airport BL Bridge - RC-Airport-1-16-1991 -- (RCFCWCD) 16 images
- 1991 Big Morongo - RC-1-34-1991 -- (RCFCWCD) 33 images
- 1991 Blythe - RC-Blythe-A-1991 -- (RCFCWCD) 1 image
- 1991 Blythe Sanitary Landfill - RC-Blythe-1-3-1991 -- (RCFCWCD) 3 images
- 1991 Cathedral Canyon East Channel - RC-East-1-16-1991 -- (RCFCWCD) 16 images
- 1991 Cathedral Canyon West Channel - RC-West-1-18-1991 -- (RCFCWCD) 18 images
- 1991 Coachella - RC-Coachella-A-1991 -- (RCFCWCD) 1 image
- 1991 Coachella Sanitary Landfill - RC-Coachella-1-21-1991 -- (RCFCWCD) 21 images
- 1991 Desert Center Sanitary Landfill - RC-Desert-A-1991 -- (RCFCWCD) 1 image
- 1991 Little Morongo - RC-61-78-1991 -- (RCFCWCD) 18 images
- 1991 Long Canyon Channel - RC-1-24-1991 -- (RCFCWCD) 24 images
- 1991 Mission Creek - RC-Mission-1-35-1991 -- (RCFCWCD) 35 images
- 1991 Mission Creek - RC-35-60-1991 -- (RCFCWCD) 26 images
- 1991 Oasis Sanitary Landfill - RC-Oasis-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Oasis Sanitary Landfill - RC-Oasis-A-1991 -- (RCFCWCD) 1 image
- 1991 Palm Canyon Channel - RC-1-35-1991 -- (RCFCWCD) 35 images
- 1991 Tahchevah Basin - RC-Tachevah-1-6-1991 -- (RCFCWCD) 6 images
- 1991 Tahquitz Debris Basin - RC-Tahquitz-1-6-1991 -- (RCFCWCD) 6 images
- 1991 Wide Canyon Basin - RC-Wide-1-8-1991 -- (RCFCWCD) 8 images
- 1992 Cathedral Canyon - RC-1-17-1992 -- (RCFCWCD) 17 images
- 1992 Cathedral Canyon West Channel - RC-1-16-1992 -- (RCFCWCD) 16 images
- 1992 Cathedral Canyon East Channel - RC-1-17-1992 -- (RCFCWCD) 17 images
- 1992 Coachella Sanitary Landfill Spot - RC-Coachella-A-1992 -- (RCFCWCD) 1 image
- 1992 Desert Center Sanitary Landfill - RC-1-3-1992 -- (RCFCWC 3 images
- 1992 Edom Hill Sanitary Landfill - RC-1-20-1992 -- (RCFCWC 19 images
- 1992 Edom Hill Sanitary Landfill - RC-Edom-A-1992-A -- (RCFCWCD) 1 image
- 1992 Mecca Sanitary Landfill - RC-1-5-1992 -- (RCFCWCD) 5 images
- 1992 Morongo Wash Stage 1 - RC-Morongo-1-26-1992 -- (RCFCWCD) 26 images
- 1992 Palm Springs Ortho Phase II - RC-71-112-1992 -- (RCFCWCD) 41 images
- 1992 Palm Springs Ortho Phase II - RC-113-150-1992 -- (RCFCWCD) 38 images
- 1992 Palm Springs Ortho Annex - RC-52-70-1992 -- (RCFCWCD) 19 images
- 1992 Palm Springs Ortho Phase 1 - RC-1-50-1992 -- (RCFCWCD) 50 images
- 1992 Riverside / San Bernardino - RC-K33-204-1992 -- (RCFCWCD) 56 images
- 1992 Tahchevah Basin - RC-Tahchevah-1-6-1992 -- (RCFCWCD) 6 images
- 1992 Taquitz Dam - RC-Taquitz-1-3-1992 -- (RCFCWCD) 3 images
- 1992 Thermal - RC-Thermal-1-10-1992 -- (RCFCWCD) 10 images
- 1992 Thermal - RC-Thermal-A-1992 -- (RCFCWCD) 1 image
- 1992 Wide Canyon Basin - RC-Wide-1-8-1992 -- (RCFCWCD) 8 images
- 1993 Black Rock - RC-Black-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Blythe Sanitary Landfill - RC-Blythe-1-4-1993 -- (RCFCWCD) 4 images
- 1993 Blythe Sanitary Landfill - RC-Blythe2-1-9-1993 -- (RCFCWCD) 4 images
- 1993 Bradshaw - RC-Bradshaw-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Coachella Spot - RC-1-22-1993 -- (RCFCWCD) 22 images
- 1993 Desert Center Sanitary Landfill - RC-1-4-1993 -- (RCFCWCD) 4 images
- 1993 Desert Hot Springs - RC-Desert-1-9-1993 -- (RCFCWCD) 9 images
- 1993 Edom Hill Sanitary Landfill - RC-Edom-1-21-1993 -- (RCFCWCD) 21 images
- 1993 Mecca Sanitary Landfill - RC-1-6-1993 -- (RCFCWCD) 6 images
- 1993 Midland - RC-Midland-1-5-1993 -- (RCFCWCD) 5 images
- 1993 Oasis Sanitary Landfill - RC-1-5-1993 -- (RCFCWCD) 5 images
- 1993 Palm Canyon - RC-1-78-1993 -- (RCFCWCD) 73 images
- 1993 Palm Canyon Wash - RC-1-30-1993 -- (RCFCWCD) 30 images
- 1993 Tahquitz Dam - RC-Tahquitz-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Thermal Canyon - RC-Thermal-1-4-1993 -- (RCFCWCD) 4 images
- 1993 Whitewater River - RC-1-13-1993 -- (RCFCWCD) 13 images
- 1993 Whitewater River - RC-14-43-1993 -- (RCFCWCD) 30 images
- 1994 Blythe Sanitary Landfill - RC-1-12-1994 -- (RCFCWCD) 11 images
- 1994 Blythe Spot - RC-Blythe-A-1994 -- (RCFCWCD) 1 image
- 1994 Coachella Sanitary Landfill - RC-1-20-1994 -- (RCFCWCD) 20 images
- 1994 Coachella Spot - RC-Coachella-A-1994 -- (RCFCWCD) 1 image
- 1994 Desert Center Sanitary Landfill - RC-1-3-1994 -- (RCFCWCD) 3 images
- 1994 Desert Center Sanitary Landfill - RC-Desert2-1-3-1994 -- (RCFCWCD) 3 images
- 1994 Desert Center Sanitary Landfill Spot - RC-1-1994 -- (RCFCWCD) 1 image
- 1994 Desert Center Spot - RC-Desert-A-1994 -- (RCFCWCD) 1 image
- 1994 Mecca Sanitary Landfill - RC-Mecca2-1-5-1994 -- (RCFCWCD) 5 images
- 1994 Mecca II Sanitary Landfill - RC-1-6-1994 -- (RCFCWCD) 6 images
- 1994 Mecca II Sanitary Landfill Spot - RC-Mecca1-1-1994 -- (RCFCWCD) 1 image
- 1994 Mecca II Spot - RC-Mecca-A-1994 -- (RCFCWCD) 1 image
- 1994 Oasis Sanitary Landfill - RC-Oasis2-1-4-9-1994 -- (RCFCWCD) 4 images
- 1994 Oasis Sanitary Landfill - RC-Oasis2-1-3-1994 -- (RCFCWCD) 3 images
- 1994 Oasis Sanitary Landfill Spot - RC-Oasis-1-1994 -- (RCFCWCD) 1 image
- 1994 Oasis Spot - RC-Oasis-A-1994 -- (RCFCWCD) 1 image
- 1994 Palm Canyon - RC-1-54-1994 -- (RCFCWCD) 54 images
- 1994 Tahquitz Dam - RC-Tahquitz-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Wide Canyon Dam - RC-Wide-1-7-1994 -- (RCFCWCD) 7 images
- 1995 Blythe Sanitary Landfill - RC-1-12-1995 -- (RCFCWCD) 12 images
- 1995 Blythe Sanitary Landfill - RC-Blythe-1-4-1995 -- (RCFCWCD) 4 images
- 1995 Blythe Sanitary Landfill Spot - RC-Blythe-A-1995 -- (RCFCWCD) 1 image
- 1995 Cathedral Canyon Channel East - RC-Cathedral-1-17-1995 -- (RCFCWCD) 17 images
- 1995 Cathedral Canyon Channel West - RC-Cathedral2-1-17-1995 -- (RCFCWCD) 17 images
- 1995 Chino Levee to Whitewater - RC-Chino-1-55-1995 -- (RCFCWCD) 55 images
- 1995 Coachella Sanitary Landfill - RC-1-18-1995 -- (RCFCWCD) 18 images
- 1995 Coachella Sanitary Landfill Spot - RC-Coachella-A-1995 -- (RCFCWCD) 1 image
- 1995 Desert Hot Springs Channel - RC-Desert-1-9-1995 -- (RCFCWCD) 9 images
- 1995 Desert Center Sanitary Landfill - RC-Desert3-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Desert Center Sanitary Landfill Spot - RC-Desert2-A-1995 -- (RCFCWCD) 1 image
- 1995 Edom Hill Sanitary Landfill - RC-1-19-1995 -- (RCFCWCD) 19 images
- 1995 Edom Hill Sanitary Landfill - RC-Edom-A-1995 -- (RCFCWCD) 1 image
- 1995 Edom Hill Sanitary Landfill - RC-Edom-B-1995 -- (RCFCWCD) 1 image
- 1995 Mission Creek Channel - RC-1-34-1995 -- (RCFCWCD) 34 images
- 1995 Oasis Sanitary Landfill - RC-Oasis2-1-4-1995 -- (RCFCWCD) 4 images
- 1995 Oasis Sanitary Landfill Spot - RC-Oasis-A-1995 -- (RCFCWCD) 1 image
- 1995 Palm Canyon Wash - RC-1-51-1995 -- (RCFCWCD) 51 images
- 1995 Palm Canyon Wash - RC-1-58-1995 -- (RCFCWCD) 58 images
- 1995 Tahchevah Basin - RC-Tahchevah-1-6-1995 -- (RCFCWCD) 6 images
- 1995 Tahquitz Basin - RC-Tahquitz-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Tahquitz Creek Debris Basin - RC-Tahquitz2-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Wide Canyon Dam - RC-Wide-1-6-1995 -- (RCFCWCD) 6 images
- 1996 Blythe Sanitary Landfill Apot - RC-Blythe-A-1996 -- (RCFCWCD) 1 image
- 1996 Blythe Sanitary Landfill - RC-Blythe2-1-12-1996 -- (RCFCWCD) 12 images
- 1996 Coachella Sanitary Landfill - RC-1-18-1996 -- (RCFCWCD) 18 images
- 1996 Coachella Sanitary Landfill Spot - RC-Coachella-A-1996 -- (RCFCWCD) 1 image
- 1996 Desert Hot Springs Ortho - RC-1-66-1996 -- (RCFCWCD) 66 images
- 1996 Edom Hill Sanitary Landfill - RC-1-21-1996 -- (RCFCWCD) 21 images
- 1996 Edom Hill Sanitary Landfill Spot - RC-Edom-A-1996 -- (RCFCWCD) 1 image
- 1996 Edom Hill Sanitary Landfill Spot - RC-Edom-B-1996 -- (RCFCWCD) 1 image
- 1996 Mecca II Sanitary Landfill - RC-Mecca2-1-5-1996 -- (RCFCWCD) 5 images
- 1996 Mecca II Sanitary Landfill Spot - RC-Mecca-A-1996-A -- (RCFCWCD) 1 image
- 1996 Palm Canyon - RC-Palm2-1-30-1996 -- (RCFCWCD) 30 images
- 1996 Palm Springs Master Drainage Plan - RC-PS-1-9-1996 -- (RCFCWCD) 9 images
- 1996 Palm Springs Main Drain Pipe Line 23 - RC-Palm-1-7-1996 -- (RCFCWCD) 7 images
- 1996 Tahquitz Dam - RC-Tahquitz-1-3-1996 -- (RCFCWCD) 3 images
- 1996 Whitewater River Levee Stage 11 - RC-1-41-1996 -- (RCFCWCD) 41 images
- 1997 Blythe - RC-Blythe-4-1-1997 -- (RCFCWCD) 6 images
- 1997 Blythe Sanitary Landfill - RC-Blythe-1-1-3-4-1997 -- (RCFCWCD) 17 images
- 1997 Coachella Sanitary Landfill - RC-1-1-3-6-1997 -- (RCFCWCD) 18 images
- 1997 Coachella Sanitary Landfill Spot - RC-Coachella-4-1-1997 -- (RCFCWCD) 1 image
- 1997 Desert Center - RC-Desert-1-3-1997 -- (RCFCWCD) 1 image
- 1997 Desert Center Sanitary Landfill - RC-1-3-1997 -- (RCFCWCD) 3 images
- 1997 Desert Center Sanitary Landfill Spot - RC-A-1997 -- (RCFCWCD) 1 image
- 1997 Desert Center Spot - RC-Desert2-2-1-1997 -- (RCFCWCD) 1 image
- 1997 Desert Hot Springs Channel - RC-Desert3-1-10-1997 -- (RCFCWCD) 10 images
- 1997 East Cathedral Channel - RC-East-1-16-1997 -- (RCFCWCD) 15 images
- 1997 Edom Hill Sanitary Landfill - RC-1-1-3-7-1997 -- (RCFCWCD) 21 images
- 1997 Edom Hill Sanitary Landfill Spot - RC-Edom-4-1-1997 -- (RCFCWCD) 1 image
- 1997 Edom Hill Sanitary Landfill Spot - RC-Edom2-5-1-1997 -- (RCFCWCD) 1 image
- 1997 Mecca II Sanitary Landfill - RC-Mecca2-1-5-1997 -- (RCFCWCD) 5 images
- 1997 Mecca II Sanitary Landfill - RC-1-5-1997 -- (RCFCWCD) 5 images
- 1997 Mecca II Sanitary Landfill Spot - RC-Mecca3-2-1-1997 -- (RCFCWCD) 1 image
- 1997 Mecca II Sanitary Landfill Spot - RC-Mecca-A-1997 -- (RCFCWCD) 1 image
- 1997 Mission Creek - RC-1-37-1997 -- (RCFCWCD) 37 images
- 1997 Oasis Sanitary Landfill - RC-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Oasis Sanitary Landfill - RC-Oasis-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Oasis Sanitary Landfill Spot - RC-Oasis2-2-1-1997 -- (RCFCWCD) 1 image
- 1997 Palm Canyon Wash - RC-1-58-1997 -- (RCFCWCD) 51 images
- 1997 Tahchevah Basin - RC-Tahchevah-1-5-1997 -- (RCFCWCD) 5 images
- 1997 Tahquitz Dam - RC-Tahquitz-1-3-1997 -- (RCFCWCD) 3 images
- 1997 West Cathedral Channel - RC-West-1-16-1997 -- (RCFCWCD) 16 images
- 1997 Whitewater River Level - RC-1-47-1997 -- (RCFCWCD) 47 images
- 1997 Wide Canyon Basin - RC-1-10-1997 -- (RCFCWCD) 10 images
- 1998 Coachella Sanitary Landfill Spot - RC-Coachella-A-1998 -- (RCFCWCD) 1 image
- 1998 Coachella Sanitary Landfill - RC-Coachella2-1-19-1998 -- (RCFCWCD) 19 images
- 1998 Desert Center Sanitary Landfill - RC-Desert2-1-3-1998 -- (RCFCWCD) 3 images
- 1998 Desert Center Sanitary Landfill Spot - RC-Desert-1998 -- (RCFCWCD) 1 image
- 1998 Desert Hot Springs Channel - RC-Desert-1-9-1998 -- (RCFCWCD) 9 images
- 1998 Eagle Canyon Dam - RC-Eagle-1-5-1998 -- (RCFCWCD) 5 images
- 1998 Mecca II Sanitary Landfill - RC-1-5-1998 -- (RCFCWCD) 5 images
- 1998 Mecca II Sanitary Landfill Spot - RC-Mecca-1998 -- (RCFCWCD) 1 image
- 1998 Oasis Sanitary Landfill - RC-Oasis2-1-4-1998 -- (RCFCWCD) 4 images
- 1998 Oasis Sanitary Landfill Spot - RC-Oasis-1998 -- (RCFCWCD) 1 image
- 1998 Palm Spring Main Drain Pipe Line 41 - RC-Palm-1-6-1998 -- (RCFCWCD) 6 images
- 1998 Tehchevah Basin - RC-1-6-1998 -- (RCFCWCD) 6 images
- 1998 Tahquitz Dam - RC-Tahquitz2-1-3-1998 -- (RCFCWCD) 3 images
- 1998 Tahquitz Dam - RC-Tahquitz-1-3-1998 -- (RCFCWCD) 3 images
- 1998 Tahchevah Basin - RC-Tahchevah-1-6-1998 -- (RCFCWCD) 6 images
- 1999 Blythe Sanitary Landfill - RC-15-1999 -- (RCFCWCD) 15 images
- 1999 Blythe Sanitary Landfill Spot - RC-Blythe-A-1999 -- (RCFCWCD) 1 image
- 1999 Coachella Sanitary Landfill - RC-1-18-1999 -- (RCFCWCD) 18 images
- 1999 Coachella Sanitary Landfill Spot - RC-Coachella-A-1999 -- (RCFCWCD) 1 image
- 1999 East Cathedral Canyon Channel - RC-1-18-1999 -- (RCFCWCD) 18 images
- 1999 Edom Hill Sanitary Landfill Spot - RC-Edom-A-1999 -- (RCFCWCD) 1 image
- 1999 Mecca II Sanitary Landfill - RC-1-5-1999 -- (RCFCWCD) 5 images
- 1999 Mission Creek - RC-1-39-1999 -- (RCFCWCD) 39 images
- 1999 Mission Creek Channel - RC-Mission-1-43-1999 -- (RCFCWCD) 43 images
- 1999 Oasis Sanitary Landfill - RC-1-4-1999 -- (RCFCWCD) 4 images
- 1999 Palm Canyon Channel - RC-1-34-1999 -- (RCFCWCD) 34 images
- 1999 West Cathedral Canyon Channel - RC-West-1-16-1999 -- (RCFCWCD) 16 images
- 1999 Whitewater - RC-1-55-1999 -- (RCFCWCD) 55 images
- 1999 Whitewater River - RC-Whitewater-1-55-1998 -- (RCFCWCD) 55 images
- 1999 Wide Canyon Dam - RC-1-13-1998 -- (RCFCWCD) 13 images
- 2000 Blythe Sanitary Landfill - RC-Blythe2-1-12-2000 -- (RCFCWCD) 12 images
- 2000 Blythe Sanitary Landfill Spot - RC-Blythe-A-2000 -- (RCFCWCD) 1 image
- 2000 Coachella Sanitary Landfill Spot - RC-Coachella-A-2000 -- (RCFCWCD) 1 image
- 2000 Desert Center - RC-1-3-2000 -- (RCFCWCD) 3 images
- 2000 Desert Hot Springs Channel - RC-1-9-2000-- (RCFCWCD) 9 images
- 2000 Desert Hot Springs Channel - RC-Desert-1-9-2000 -- (RCFCWCD) 9 images
- 2000 East Cathedral Canyon Channel - RC-1-16-2000 -- (RCFCWCD) 17 images
- 2000 East Cathedral Canyon Channel - RC-East-1-18-2000 -- (RCFCWCD) 18 images
- 2000 Mecca II Sanitary Landfill - RC-1-5-2000 -- (RCFCWCD) 5 images
- 2000 Mecca II Sanitary Landfill Spot - RC-A-2000 -- (RCFCWCD) 1 image
- 2000 Mission Creek - RC-1-42-2000 -- (RCFCWCD) 43 images
- 2000 Oasis Sanitary Landfill - RC-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Palm Canyon - RC-Palm-1-35-2000 -- (RCFCWCD) 35 images
- 2000 Palm Canyon Channel - RC-1-35-2000 -- (RCFCWCD) 35 images
- 2000 Tahchevah Basin - RC-1-6-2000 -- (RCFCWCD) 6 images
- 2000 Tahchevah Dam - RC-Tahchevah-1-6-2000 -- (RCFCWCD) 6 images
- 2000 Tahquitz Creek Dam - RC-Tahquitz-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Tahquitz Dam - RC-Tahquitz-1-3-2000 -- (RCFCWCD) 3 images
- 2000 West Cathedral Canyon Channel - RC-1-17-2000 -- (RCFCWCD) 17 images
- 2000 West Cathedral Canyon Channel - -RC-West-1-16-2000 -- (RCFCWCD) 16 images
- 2000 White Water - RC-1-54-2000 -- (RCFCWCD) 54 images
- 2000 Wide Canyon Dam - RC-1-14-2000 -- (RCFCWCD) 14 images
- 2000 Wide Canyon Dam - RC-Wide-1-14-2000 -- (RCFCWCD) 14 images
- 2001 Blythe Sanitary Landfill - RC-1-15-2001 -- (RCFCWCD) 15 images
- 2001 Desert Hot Springs - RC-1-9-2001 -- (RCFCWCD) 9 images
- 2001 Eagle Canyon Dam - RC-1-4-2001 -- (RCFCWCD) 4 images
- 2001 East Cathedral Canyon Channel - RC-East-1-17-2001 -- (RCFCWCD) 17 images
- 2001 Mission Creek Channel - RC-1-39-2001 -- (RCFCWCD) 39 images
- 2001 Palm Canyon - RC-1-36-2001 -- (RCFCWCD) 36 images
- 2001 Tahchevah Basin - RC-Tahchevah-RC-1-6-2001 -- (RCFCWCD) 6 images
- 2001 Tahquitz Dam - RC-Tahquitz-1-3-2001 -- (RCFCWCD) 3 images
- 2001 West Cathedral Canyon Channel - RC-West-1-19-2001 -- (RCFCWCD) 19 images
- 2001 Whitewater River Right Levee - RC-1-53-2001 -- (RCFCWCD) 53 images
- 2001 Wide Canyon Basin - RC-Wide-1-12-2001 -- (RCFCWCD) 12 images
- 2002 Chino Canyon Levee - RC-1-29-2002 -- (RCFCWCD) 29 images
- 2002 East Cathedral Canyon - RC-East-1-10-2002 -- (RCFCWCD) 16 images
- 2002 Palm Canyon Cathdral - RC-1-27-2002 -- (RCFCWCD) 27 images
- 2002 Palm Canyon Wash Lateral A - RC-Palm-1-7-2002 -- (RCFCWCD) 7 images
- 2002 Tahquitz Creek Basin - RC-Tahquitz-1-4-2002 -- (RCFCWCD) 4 images
- 2002 Whitewater River - RC-1-39-2002 -- (RCFCWCD) 39 images
- 2003 Blythe Sanitary Landfill - RC-Blythe-12-15-2003 -- (RCFCWCD) 15 images
- 2003 Desert Hot Springs - RC-Desert-1-5-2003 -- (RCFCWCD) 5 images
- 2003 Edom Hill Sanitary Landfill - RC-1-22-2003 -- (RCFCWCD) 22 images
- 2003 Farrel Basin - RC-Farrel-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Farrel Basin - RC-Farrel-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Mecca II - RC-1-5-2003 -- (RCFCWCD) 5 images
- 2003 Mecca II Sanitary Landfill - RC-Mecca2-1-5-2003 -- (RCFCWCD) 5 images
- 2003 Mecca II Sanitary Landfill Spot - RC-Mecca-A-2003 -- (RCFCWCD) 1 image
- 2003 Mission Creek Channel - RC-1-30-2003 -- (RCFCWCD) 30 images
- 2003 Tahchevah Dam - RC-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Tahchevah Dam - RC-Tahchevah-1-6-2003 -- (RCFCWCD) 6 images
- 2003 Tahquitz Creek Basin - RC-Tahquitz-1-4-2003 -- (RCFCWCD) 4 images
- 2003 West Cathedral Canyon - RC-1-14-2003 -- (RCFCWCD) 14 images
- 2003 West Cathedral Canyon - RC-1-15-2003 -- (RCFCWCD) 15 images
- 2003 Wide Canyon Dam - RC-Wide2-1-9-2003 -- (RCFCWCD) 9 images
- 2003 Wide Canyon Dam - RC-Wide-1-9-2003 -- (RCFCWCD) 9 images
- 2004 Blythe Sanitary Landfill - RC-Blythe-1-15-2004 -- (RCFCWCD) 15 images
- 2004 Chino Canyon Levee - RC-1-4-2004 -- (RCFCWCD) 4 images
- 2004 Chino Canyon Levee- RC-5-26-2004 -- (RCFCWCD) 22 images
- 2004 Chino Canyon Levee - RC-1-24-2004 -- (RCFCWCD) 24 images
- 2004 Desert Hot Springs - RC-Desert-1-5-2004 -- (RCFCWCD) 5 images
- 2004 Desert Hot Springs Channel - RC-Desert-1-5-2004 -- (RCFCWCD) 5 images
- 2004 East Cathedral Canyon - RC-1-20-2004 -- (RCFCWCD) 20 images
- 2004 Ferrell Basin - RC-Ferrell-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Mission Lake Levee - RC-1-7-2004 -- (RCFCWCD) 7 images
- 2004 Mission Creek Channel - RC-1-29-2004 -- (RCFCWCD) 29 images
- 2004 North Palm Springs - RC-1-5-2004 -- (RCFCWCD) 5 images
- 2004 Palm Springs Ortho Phase II - RC-Palm-1-37-2004 -- (RCFCWCD) 37 images
- 2004 Palm Springs Ortho Phase III - RC-Palm2-1-37-2004 -- (RCFCWCD) 37 images
- 2004 Palm Canyon Channel - RC-1-26-2004 -- (RCFCWCD) 26 images
- 2004 Palm Canyon Wash Lateral A - RC-Palm-1-7-2004 -- (RCFCWCD) 7 images
- 2004 Tahchevah Dam - RC-Tahchevah-1-6-2004 -- (RCFCWCD) 6 images
- 2004 Tahquitz Creek Basin - RC-Tahquitz-1-4-2004 -- (RCFCWCD) 4 images
- 2004 Verbena Channel - RC-1-29-2004 -- (RCFCWCD) 28 images
- 2004 Whitewater River - RC-1-41-2004 -- (RCFCWCD) 41 images
- 2004 Whitewater River - RC-1-38-2004 -- (RCFCWCD 38 images
- 2004 Wide Canyon Dam - RC-Wide-1-9-2004 -- (RCFCWCD) 9 images
- 2005 East Cathedral Canyon - RC-1-14-2005 -- (RCFCWCD) 14 images
- 2005 Edom Hill Sanitary Landfill - RC-1-31-2005 -- (RCFCWCD) 31 images
- 2005 Mecca II Sanitary Landfill - RC-A-2005 -- (RCFCWCD) 1 image
- 2005 Mecca II Sanitary Landfill - RC-Mecca-1-5-2005 -- (RCFCWCD) 5 images
- 2005 Mission Creek Channel - RC-1-29-2005 -- (RCFCWCD) 29 images
- 2005 North Palm Springs Ortho - RC-1-33-2005 -- (RCFCWCD) 33 images
- 2005 Palm Canyon Channel - RC-1-26-2005 -- (RCFCWCD) 26 images
- 2005 Palm Canyon Wash - RC-1-16-2005 -- (RCFCWCD) 16 images
- 2005 Palm Canyon Wash Lateral A - RC-1-7-2005 -- (RCFCWCD) 7 images
- 2005 Tahquitz Creek Basin - RC-Tahquitz-1-4-2005 -- (RCFCWCD) 4 images
- 2005 West Cathedral Canyon - RC-1-13-2005 -- (RCFCWCD) 13 images
- 2006 Chino Canyon Levee - RC-Chino-1-25-2006 -- (RCFCWCD) 25 images
- 2006 Chino Canyon Levee - RC-Chino6-1-25-2006 -- (RCFCWCD) 25 images
- 2006 Desert Hot Springs Channel - RC-Desert-1-5-2006 -- (RCFCWCD) 5 images
- 2006 Desert Hot Springs Channel - RC-Desert6-1-5-2006 -- (RCFCWCD) 5 images
- 2006 East Cathedral Canyon - RC-1-14-2006 -- (RCFCWCD) 14 images
- 2006 East Cathedral Canyon - RC-E-1-14-2006 -- (RCFCWCD) 14 images
- 2006 Ferrell Basin - RC-Ferrell-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Ferrell Basin - RC-Ferrell6-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Mission Creek Channel - RC-1-29-2006 -- (RCFCWCD) 29 images
- 2006 Morongo Basin Area - RC-1-36-2006 -- (RCFCWCD) 36 images
- 2006 Palm Canyon Channel - RC-Palm-1-25-2006 -- (RCFCWCD) 25 images
- 2006 Palm Canyon Channel - RC-Palm6-1-26-2006 -- (RCFCWCD) 26 images
- 2006 Palm Canyon Wash - RC-1-7-2006 -- (RCFCWCD) 7 images
- 2006 Palm Canyon Wash Lateral A - RC-Palm6-1-8-2006 -- (RCFCWCD) 8 images
- 2006 Tahchevah Dam - RC-Tahchevah-1-6-2006 -- (RCFCWCD) 6 images
- 2006 Tahchevah Dam - RC-Tahchevah6-1-14-2006 -- (RCFCWCD) 11 images
- 2006 Tahquitz Creek Basin - RC-1-4-2006 -- (RCFCWCD) 4 images
- 2006 Tahquitz Creek Basin - RC-Tahquitz6-1-4-2006 -- (RCFCWCD) 4 images
- 2006 West Cathedral Canyon - RC-West-1-14-2006 -- (RCFCWCD) 14 images
- 2006 West Cathedral Canyon - RC-W-1-10-2006 -- (RCFCWCD) 10 images
- 2006 Whitewater River - RC-Whitewater-1-37-2006 -- (RCFCWCD) 37 images
- 2006 Whitewater River - RC-1-39-2006 -- (RCFCWCD) 40 images
- 2006 Wide Canyon Dam - RC-1-9-2006 -- (RCFCWCD) 9 images
- 2006 Wide Canyon Dam - RC-Wide6-1-9-2006 -- (RCFCWCD) 9 images
- 2007 East Cathedral Canyon - RC-East2-1-15-2007-- (RCFCWCD) 15 images
- 2007 Palm Canyon Channel - RC-1-35-2007-- (RCFCWCD) 35 images
- 2007 West Cathedral Canyon - RC-West2-1-13-2007-- (RCFCWCD) 13 images
- 2007 Whitewater River - RC-1-7-2007-- (RCFCWCD) 7 images
- 2008 Chino Canyon Levee - RC-1-26-2008 -- (RCFCWCD) 26 images
- 2008 Desert Hot Springs Channel - RC-Desert7-1-5-2008 -- (RCFCWCD) 5 images
- 2008 Desert Hot Springs Channel - RC-Desert-1-5-2008 -- (RCFCWCD) 5 images
- 2008 East Cathedral Canyon - RC-East7-1-15-2008 -- (RCFCWCD) 15 images
- 2008 Edom Hill Sanitary Landfill - RC-Edom-1-30-2008 -- (RCFCWCD) 30 images
- 2008 Farrel Basin - RC-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Ferrell Basin - RC-Ferrell7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Hemet Channel - RC-1-24-2008 -- (RCFCWCD) 24 images
- 2008 Mission Creek Channel - RC-Mission-1-29-2008 -- (RCFCWCD) 29 images
- 2008 Mission Creek Channel - RC-1-29-2008 -- (RCFCWCD) 28 images
- 2008 Morongo Basin - RC-1-36-2008 -- (RCFCWCD) 36 images
- 2008 Palm Canyon Channel - RC-1-35-2008 -- (RCFCWCD) 35 images
- 2008 Palm Canyon Wash - RC-1-7-2008 -- (RCFCWCD) 7 images
- 2008 Palm Canyon Wash Lateral A - RC-Palm7-1-7-2008 -- (RCFCWCD) 7 images
- 2008 San Jacinto River Stage IV Extension - RC-San7-1-15-2008 -- (RCFCWCD) 15 images
- 2008 Tahchevah Dam - RC-Tahchevah-1-6-2008 -- (RCFCWCD) 6 images
- 2008 Tahchevah Dam - RC-Tahchevah7-1-6-2008 -- (RCFCWCD) 6 images
- 2008 Tahquitz Creek Basin - RC-Tahquitz-1-4-2008 -- (RCFCWCD) 4 images
- 2008 Tahquitz Creek Basin - RC-Tahquitz7-1-3-2008 -- (RCFCWCD) 3 images
- 2008 West Cathedral Canyon - RC-West8-1-13-2008 -- (RCFCWCD) 14 images
- 2008 Whitewater River - RC-8-51-2008 -- (RCFCWCD) 44 images
- 2008 Whitewater River - RC-Whitewater7-1-7-2008 -- (RCFCWCD) 7 images
- 2008 Wide Canyon Dam - RC-Wide8-1-9-2008 -- (RCFCWCD) 9 images
- 2008 Wide Canyon Dam - RC-Wide7-1-9-2008 -- (RCFCWCD) 9 images
- 2009 Chino Canyon Levee - RC-Chino7-1-25-2009 -- (RCFCWCD) 25 images
- 2009 Desert Hot Springs Channel - RC-Desert7-1-5-2009 -- (RCFCWCD) 5 images
- 2009 Farrel Basin - RC-Farrel-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Long Canyon Ortho - RC-1-104-2009 -- (RCFCWCD) 104 images
- 2009 Mission Creek Channel - RC-1-29-2009 -- (RCFCWCD) 29 images
- 2009 Palm Canyon Channel - RC-1-35-2009 -- (RCFCWCD) 35 images
- 2009 Palm Canyon Wash Lateral A - RC-Palm7-1-7-2009 -- (RCFCWCD) 7 images
- 2009 Palm Springs Line 43 & 43A - RC-PS-1-8-2009 -- (RCFCWCD) 8 images
- 2009 Tehchevah Dam - RC-Tehchevah7-1-6-2009 -- (RCFCWCD) 6 images
- 2009 Whitewater River - RC-1-51-2009 -- (RCFCWCD) 51 images
- 2009 Wide Canyon Dam - RC-Wide7-1-9-2009 -- (RCFCWCD) 9 images
- 2011 Blythe Sanitary Landfill - RC-1-17-2011 -- (RCFCWCD) 17 images
- 2011 Blythe Sanitary Landfill Spot - RC-A-2011 -- (RCFCWCD) 1 image
- 2011 Chino Canyon Levee - RC-1-25-2011 -- (RCFCWCD) 25 images
- 2011 Coachella Sanitary Landfill - RC-1-18-2011 -- (RCFCWCD) 18 images
- 2011 Coachella Sanitary Landfill Spot - RC-Spot-A-2011 -- (RCFCWCD) 1 image
- 2011 Desert Hot Springs Channel - RC-Desert-1-5-2011 -- (RCFCWCD) 5 images
- 2011 East Cathedral Canyon - RC-East-1-15-2011 -- (RCFCWCD) 15 images
- 2011 Farrell Basin - RC-Farrell-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Mission Creek Channel - RC-Mission-1-29-2011 -- (RCFCWCD) 29 images
- 2011 Oasis Sanitary Landfill - RC-Oasis-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Palm Canyon Channel - RC-1-35-2011 -- (RCFCWCD) 35 images
- 2011 Palm Canyon Wash Lateral A - RC-Palm-1-9-2011 -- (RCFCWCD) 9 images
- 2011 Tahchevah Dam - RC-Tahchevah-1-6-2011 -- (RCFCWCD) 6 images
- 2011 Tahquitz Creek Basin - RC-Tahquitz-1-8-2011 -- (RCFCWCD) 8 images
- 2011 West Cathedral Canyon - RC-1-17-2011 -- (RCFCWCD) 17 images
- 2011 Whitewater River - RC-1-51-2011 -- (RCFCWCD) 51 images
- 2011 Wide Canyon Dam- RC-1-9-2011 -- (RCFCWCD) 9 images
- 2012 Canyon Springs Basin - RC-Canyon-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Chino Canyon Levee - RC-Chino7-1-25-2012 -- (RCFCWCD) 25 images
- 2012 Desert Hot Springs Channel - RC-Desert-1-5-2012 -- (RCFCWCD) 5 images
- 2012 Eagle Canyon Dam - RC-Eagle7-1-5-2012 -- (RCFCWCD) 5 images
- 2012 East Cathedral Canyon - RC-1-15-2012 -- (RCFCWCD) 15 images
- 2012 Farrell Basin - RC-Farrell-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Mission Creek Channel - RC-1-29-2012 -- (RCFCWCD) 29 images
- 2012 Palm Canyon Channel - RC-1-35-2012 -- (RCFCWCD) 35 images
- 2012 Palm Canyon Wash Lateral A - RC-Palm7-1-7-2012 -- (RCFCWCD) 7 images
- 2012 Tahchevah Dam - RC-Tahchevah-1-6-2012 -- (RCFCWCD) 6 images
- 2012 Tahquitz Creek Basin - RC-Tahquitz7-1-6-2012 -- (RCFCWCD) 6 images
- 2012 Towngate Basin - RC-Towngate-1-2-2012 -- (RCFCWCD) 2 images
- 2012 West Cathedral Canyon - RC-West-1-13-2012 -- (RCFCWCD) 13 images
- 2012 Whitewater River - RC-1-51-2012 -- (RCFCWCD) 51 images
- 2012 Wide Canyon Dam - RC-Wide-1-9-2012 -- (RCFCWCD) 9 images
- 2013 Chino Canyon Levee - RC-Chino-1-25-2013 -- (RCFCWCD) 25 images
- 2013 Desert Hot Springs Channel - RC-Hot-1-5-2013 -- (RCFCWCD) 5 images
- 2013 Desert Recycling - RC-Desert-1-5-2013 -- (RCFCWCD) 5 images
- 2013 East Cathedral Canyon- RC-East-1-15-2013 -- (RCFCWCD) 15 images
- 2013 Farrell Basin - RC-Farrell-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Mission Creek Channel - RC-1-29-2013 -- (RCFCWCD) 29 images
- 2013 Palm Canyon Channel - RC-1-35-2013 -- (RCFCWCD) 35 images
- 2013 Palm Canyon Wash Lateral A - RC-Palm-1-7-2013 -- (RCFCWCD) 7 images
- 2013 Tahchevah Dam - RC-Tahchevah-1-6-2013 -- (RCFCWCD) 6 images
- 2013 Tahquitz Creek Basin - RC-Tahquitz-1-6-2013 -- (RCFCWCD) 6 images
- 2013 West Cathedral Canyon - RC-West2-1-13-2013 -- (RCFCWCD) 13 images
- 2013 Whitewater River - RC-1-51-2013 -- (RCFCWCD) 51 images
- 2013 Wide Canyon Dam- RC-Wide-1-9-2013 -- (RCFCWCD) 9 images
- 2014 Edom Hills Sanitary Landfill - RC-1-30-2014 -- (RCFCWCD) 30 images
- 2014 Edom Hills Sanitary Landfill Spot - RC-Edom-A-2014 -- (RCFCWCD) 1 image
- 2015 Chino Canyon Levee - RC-1-25-2015 -- (RCFCWCD) 25 images
- 2015 Desert Hot Springs Channel - RC-Desert-1-5-2015 -- (RCFCWCD) 5 images
- 2015 East Cathedral Canyon - RC-East-1-15-2015 -- (RCFCWCD) 15 images
- 2015 Farrell Basin - RC-Farrell-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Mission Creek Channel - RC-1-29-2015 -- (RCFCWCD) 29 images
- 2015 Palm Canyon Channel - RC-1-35-2015 -- (RCFCWCD) 35 images
- 2015 Palm Canyon Wash- RC-Palm-1-15-2015 -- (RCFCWCD) 15 images
- 2015 Tahchevah Dam - RC-Tahchevah-1-6-2015 -- (RCFCWCD) 6 images
- 2015 Tahquitz Creek Basin - RC-Tahquitz-1-6-2015 -- (RCFCWCD) 6 images
- 2015 West Cathedral Canyon - RC-1-13-2015 -- (RCFCWCD) 13 images
- 2015 Whitewater River - RC-1-51-2015 -- (RCFCWCD) 51 images
- 2015 Wide Canyon Dam - RC-1-9-2015 -- (RCFCWCD) 9 images
- (Date Unknown) Salton Sea -- (Rowe) 5 images
-
Riverside-City (8,858 images)
The City of Riverside is located in Inland Southern California in the northwest portion of Riverside County.
- 1931 City of Riverside -- Flight C1740 (Rowe) 94 images
- 1938 Santa Ana River - RC-1-14-1938 -- (RCFCWCD) 14 images
- 1938 Santa Ana River - RC-AXL-AXM-35-58-62-1938 -- (RCFCWCD) 5 images
- 1938 Santa Ana River - RC-AXL-AXM-39-39-1938 -- (RCFCWCD) 1 image
- 1938 Santa Ana River - RC-AXL-AXM-40-24-72-1938 -- (RCFCWCD) 10 images
- 1938 Santa Ana River - RC-AXL-AXM-41-65-81-1938 -- (RCFCWCD) 7 images
- 1938 Santa Ana River - RC-AXL-AXM-45-44-54-1938 -- (RCFCWCD) 6 images
- 1938 Santa Ana River - RC-AXL-AXM-46-48-71-1938 -- (RCFCWCD 4 images
- 1938 Santa Ana River - RC-AXL-AXM-48-55-57-1938 -- (RCFCWCD) 3 images
- 1938 Santa Ana River - RC-AXL-AXM-54-43-43-1938 -- (RCFCWCD 1 image
- 1938 Santa Ana River - RC-AXL-AXM-61-87-89-1938 -- (RCFCWCD) 2 images
- 1938 Santa Ana River - RC-AXL-AXM-62-7-29-1938 -- (RCFCWCD) 7 images
- 1938 Santa Ana River - RC-AXL-AXM-65-8-16-1938 -- (RCFCWCD) 3 images
- 1938 Santa Ana River - RC-AXL-AXM-82-28-29-1938 -- (RCFCWCD) 2 images
- 1939 Santa Ana River - RC-AXL-AXM-209-86-86-1939 -- (RCFCWCD) 1 images
- 1947 Riverside - RC-1-6-1947 -- (RCFCWCD) 3 images
- 1948 North of County Line - RC-12-15-1948 -- (RCFCWCD) 4 images
- 1948 Riverside / Colton- RC-1-42-1948 -- (RCFCWCD) 34 images
- 1948 Riverside Santa Ana River - RC-43-78-1949 -- (RCFCWCD) 34 images
- 1949 Box Springs - RC-1-3-1949 -- (RCFCWCD) 3 images
- 1949 Mira Loma - RC-1-50-1949 -- (RCFCWCD) 9 images
- 1949 Riverside Book 3A - RC-AXM-F-3F-142-189-1949 -- (RCFCWCD) 9 images
- 1949 Riverside Book 3A - RC-AXM-F-5F-32-190-1949 -- (RCFCWCD) 24 images
- 1949 Riverside Book 3A - RC-AXM-F-6F-29-121-1949 -- (RCFCWCD) 13 images
- 1949 Riverside Book 3A - RC-AXM-F-9F-101-179-1949 -- (RCFCWCD) 13 images
- 1949 Riverside Book 3A - RC-AXM-F-10F-36-194-1949 -- (RCFCWCD) 17 images
- 1952 East Riverside Woodcrest - RC-5-3-76-1952 -- (RCFCWCD) 74 images
- 1952 Tri-City Rock Company - Flight GS-VR-1952 -- Rowe 9 images
- 1953 Riverside Book 1B - RC-AXM-6K-10-50-1953 -- (RCFCWCD) 2 images
- 1954 Riverside Book 1B - RC-AXM-16K-190-190-1954 -- (RCFCWCD) 1 image
- 1955 Mira Loma - RC-1-4 -- (RCFCWCD) 3 images
- 1957 La Sierra - RC-41-48 -- (RCFCWCD) 8 images
- 1957 University High Grove - RC-1-33 -- (RCFCWCD) 33 images
- 1958 Arlington Ave -RC-25-69 -- (RCFCWCD) 44 images
- 1958 Glen Avon - Rubidoux -RC-1-33 -- (RCFCWCD) 33 images
- 1958 West Riverside - RC-2-4 -- (RCFCWCD) 3 images
- 1959 County Farm - RC-32-34 -- (RCFCWCD) 5 images
- 1959 Home Gardens, La Sierra, Arlington - RC-1-23 -- (RCFCWCD) 23 images
- 1959 La Sierra Convergence - RC-15-51 -- (RCFCWCD) 35 images
- 1959 Limonite Ave - RC-33-65 -- (RCFCWCD) 33 images
- 1959 Pidgeon Pass, Highgrove, Reche Canyon - RC-24-38 -- (RCFCWCD) 38 images
- 1960 Arlanza - RC-37-43 -- (RCFCWCD) 7 images
- 1960 Glen Avon Heights - RC-46-50-1960 -- (RCFCWCD) 5 images
- 1960 Glen Avon Heights -RC-50-57-1960 -- (RCFCWCD) 9 images
- 1960 La Sierra - RC-26-36 -- (RCFCWCD) 10 images
- 1960 La Sierra - RC-44-46-1960 -- (RCFCWCD) 3 images
- 1960 Mira Loma - RC-26-45-1960 -- (RCFCWCD) 41 images
- 1960 Riverside County Flood Control Compound - RC-1-3-1960 -- (RCFCD) 3 images
- 1961 Central Ave Extension - RC-15-30 -- (RCFCWCD) 16 images
- 1961 La Sierra - RC-1-13 -- (RCFCWCD) 13 images
- 1961 University Highgrove - RC-1-33 -- (RCFCWCD) 2 images
- 1962 Victoria Storm Drain / Gage Canal- RC-23-51 -- (RCFCWCD) 29 images
- 1963 Riverside - RC-1-122-1963 -- (RCFCWCD) 122 images
- 1963 Santa Ana River San Sevaine - RC-123-147-1963 -- (RCFCWCD) 25 images
- 1965 La Sierra - RC-1-27-1964 -- (RCFCWCD) 27 images
- 1965 Riverside - RC-1-50-1965 -- (RCFCWCD) 53 images
- 1965 Santa Ana River Flood Damage - RC-1-8-1965 -- (RCFCWCD) 8 images
- 1965 West Riverside -RC-57-92-1965 -- (RCFCWCD) 34 images
- 1966 Highgrove - RC-1-3-1966 -- (RCFCWCD) 3 images
- 1966 Santa Ana River Outfall - RC-1-1-1-190-1966 -- (RCFCWCD) 189 images
- 1966 Santa Ana River Outfall - RC-2-1-2-51-1966 -- (RCFCWCD) 43 images
- 1966 Santa Ana River Outfall Stage II Convergents - RC45-94-1966 -- (RCFCWCD) 50 images
- 1966 Santa Ana River Rubidoux - RC-1-6-1966 -- (RCFCD) 5 images
- 1966 Sierra Ave - RC-13-25-1966 -- (RCFCWCD) 13 images
- 1966 Tri-City Rock Company - Flight GS-VBDF-1966 -- Rowe 8 images
- 1967 Arlington Storm Channel - RC-14-40-1967 -- (RCFCWCD) 27 images
- 1967 Jurupa Road - RC-1-2-1967 -- (RCFCWCD) 2 images
- 1967 La Sierra Hills - RC-12-34-1967 -- (RCFCWCD) 22 images
- 1967 La Sierra Vertical - RC-A-1967 -- (RCFCWCD) 1 image
- 1967 San Sevaine Channel - RC-1A-4A-1967 -- (RCFCWCD) 8 images
- 1967 Santa Ana River Outfall Stage III - RC-1-34-1967 -- (RCFCWCD) 34 images
- 1967 Sunnyslope - RC-1-7-2-15-1967 -- (RCFCWCD) 7 images
- 1967 Sunnyslope Extension - RC-5-12-1967 -- (RCFCWCD) 8 images
- 1968 Central Ave - RC-6-17-1968 -- (RCFCWCD) 12 images
- 1968 Merril Ave - RC-1-3-1968 -- (RCFCWCD) 3 images
- 1968 Riverside - RC-21-57-1968 -- (RCFCWCD) 37 images
- 1968 Rubidoux-Mission Blvd. - RC-1-1-1-12-1968 -- (RCFCWCD) 12 images
- 1969 Riverside Reservoirs - RC-RiversideReservoirs-1-8-1969 -- (RCFCWCD) 8 images
- 1969 Center Street -RC-1-3-1969 -- (RCFCWCD) 3 images
- 1969 Day Creek - RC-59-76-1969 -- (RCFCWCD) 18 images
- 1969 Day Creek - RC-89-100-1969 -- (RCFCWCD) 11 images
- 1969 San Sevaine - RC-1-58-1969 -- (RCFCWCD) 58 images
- 1969 San Sevaine Channel - RC-17-50-1969 -- (RCFCWCD) 34 images
- 1969 San Sevaine -RC-78-88-1969 -- (RCFCWCD) 10 images
- 1969 Santa Ana River - RC-30-64-1969 -- (RCFCWCD) 34 images
- 1969 Santa Ana River - RC-190-206-1969 -- (RCFCWCD) 17 images
- 1970 Alessandro Boulevard - RC-1-30-1970 -- (RCFCWCD) 30 imgesa
- 1970 Limonite - RC-31-65-1970 -- (RCFCWCD) 25 imgesa
- 1970 Mission Blvd - RC-1-9-1970 -- (RCFCWCD) 9 images
- 1970 Pedley - RC-5-12-1970 -- (RCFCWCD) 15 images
- 1970 Santa Ana River - RC-SAR-1-3-1970 -- (RCFCWCD) 3 images
- 1970 South East Riverside - RC-17-47-1970 -- (RCFCWCD) 30 images
- 1971 Alessandro Boulevard - RC-1-45-1971 -- (RCFCWCD) 45 images
- 1971 Archibald & Chandler - RC-1-1971 -- (RCFCWCD) 1 image
- 1971 Box Springs Project 6 - RC-1-3-1971 -- (RCFCWCD) 3 images
- 1971 Brockton Avenue - RC-1-24-1971 -- (RCFCWCD) 24 images
- 1971 Casa Blanca Area - RC-1-1971 -- (RCFCWCD) 1 image
- 1971 Central Riverside - RC-1-50-1971 -- (RCFCWCD) 50 images
- 1971 County Farm - RC-3-9-1971 -- (RCFCWCD) 7 images
- 1971 County Farm Convergence - RC-1-2-1971 -- (RCFCWCD) 2 images
- 1971 Galena Strret - RC-Galena-1-17-1971 -- (RCFCWCD) 17 images
- 1971 Jones Street Project 22 - RC-1-29-1971 -- (RCFCWCD) 29 images
- 1971 Jurupa Road - RC-1-36-1971 -- (RCFCWCD) 36 images
- 1971 Magnolia & Byron - RC-2-1971 -- (RCFCWCD) 1 image
- 1971 Magnolia Center - RC-1-1-2-8-1971 -- (RCFCWCD) 16 images
- 1971 Mary Street - RC-1-18-1971 -- (RCFCWCD) 17 images
- 1971 Mary Street - RC-12-27-1971 -- (RCFCWCD) 16 images
- 1971 Nelson Street - RC-1-11-1971 -- (RCFCWCD) 11 images
- 1971 Nelson Street - RC-19-29-1971 -- (RCFCWCD) 12 images
- 1971 Ottawa and Prince Albert - RC-1-1-1-3-1971 -- (RCFCWCD) 2 images
- 1971 Perris Reservoir - RC-1-9-1971 -- (RCFCWCD) 9 images
- 1971 Pierce Street Project 21 - RC-1-17-1971 -- (RCFCWCD) 17 images
- 1971 Rubidoux #1 - RC-1-1-1-3-1971 -- (RCFCWCD) 19 images
- 1971 Rubidoux #2 - RC-1-1-3-7-1971 -- (RCFCWCD) 26 images
- 1971 Seventh Street - RC-1-22-1971 -- (RCFCWCD) 19 images
- 1971 Tequesquite - RC-1-10-1971 -- (RCFCWCD) 10 images
- 1971 University Drain Project 6 - RC-1-35-1971 -- (RCFCWCD) 35 images
- 1972 Anza Channel - RC-1-6-1972 -- (RCFCWCD) 6 images
- 1972 Anza Channel - RC-1-35-1972 -- (RCFCWCD) 34 images
- 1972 Anza Channel - RC-1-62-1972 -- (RCFCWCD) 37 images
- 1972 Anza Channel Project 18 - RC-65-108-1972 -- (RCFCWCD) 36 images
- 1972 Anza Channel Project 18 -RC-111-144-1972 -- (RCFCWCD) 33 images
- 1972 Box Springs Project 6 - RC-1-21-1972 -- (RCFCWCD) 21 images
- 1972 Box Springs Project 6/Line C - RC-1-17-1972 -- (RCFCWCD) 17 images
- 1972 Cranford Project 7 - RC-34-49-1972 -- (RCFCWCD) 15 images
- 1972 Gramercy Place Project 20 - RC-1-41-1972 -- (RCFCWCD) 41 images
- 1972 Gramercy Place Project 20 - RC-87-110-1972 -- (RCFCWCD) 24 images
- 1972 La Sierra Avenue -RC-10-28-1972 -- (RCFCWCD) 19 images
- 1972 La Sierra Project 24 - RC-44-86-1972 -- (RCFCWCD) 42 images
- 1972 Mitchell Avenue Project 19 - RC-42-1972 -- (RCFCWCD) 1 image
- 1972 Pennsylvania Project 8 - RC-1-33-1972 -- (RCFCWCD) 33 images
- 1972 Pyrite Canyon - RC-1-9-1972 -- (RCFCWCD) 9 images
- 1973 Blair Street - Project 34 - RC-1-15-1-54-1973 -- (RCFCWCD) 22 images
- 1973 Blair Street - Project 34 - RC-1-55-1-71-1973 -- (RCFCWCD) 17 images
- 1973 Grant Street - Project 33 - RC-1-1-1-14-1973 -- (RCFCWCD) 14 images
- 1973 Grant Street - Project 33 - RC-1-28-1973 -- (RCFCWCD) 17 images
- 1973 Hole Lake Project 18 - RC-1-14-1973 -- (RCFCWCD) 14 images
- 1973 Mary Street Dam - RC-1-64-1973 -- (RCFCWCD) 64 images
- 1973 Mira Loma Revision - RC-1-58-1973 -- (RCFCWCD) 54 images
- 1973 Mira Loma Revision - RC-59-113-1973 -- (RCFCWCD) 55 images
- 1973 Monroe Storm Drain Rail Road Lateral - RC-1-1-3-3-1973 -- (RCFCWCD) 11 images
- 1973 Monroe Storm Drain Rail Road Lateral - RC-1-107-1973 -- (RCFCWCD) 105 images
- 1973 Perris Reservoir - RC-1-33-1973 -- (RCFCWCD) 31 images
- 1973 Riverside County Parks - RC-1-63-1973 -- (RCFCWCD) 53 images
- 1973 Riverside County Parks - RC-71-82-1973 -- (RCFCWCD) 12 images
- 1973 Riverside County Parks - RC-91-97-1973 -- (RCFCWCD) 7 images
- 1973 Riverside County Parks Convergence - RC-1A-7B-1973 -- (RCFCWCD) 12 images
- 1973 Riverside County Parks Convergence - RC-8A-8B-1973 -- (RCFCWCD) 2 images
- 1974 Bly Channel Mission Drain Glen Avon - RC-1-10-1974 -- (RCFCWCD) 10 images
- 1974 Etiwanda Avenue - RC-1-8-1974 -- (RCFCWCD) 8 images
- 1974 Jurupa Channel - RC-1-14-1974 -- (RCFCWCD) 24 images
- 1974 Pedley Road - RC-1-18-1974 -- (RCFCWCD) 18 images
- 1974 Riverside County Complex - RC-1-10-1974 -- (RCFCWCD) 10 images
- 1974 West Riverside Disposal (Belltown) -RC-Belltown-3-5-1974 -- (RCFCWCD) 3 images
- 1974 West Riverside Disposal Convergence - RC-1-2-1974 -- (RCFCWCD) 2 images
- 1975 Highgrove Disposal - RC-Highgrove-2-4-1975 -- (RCFCWCD) 3 images
- 1975 Mira Loma Ramp 4 - RC-27-101-1975 -- (RCFCWCD) 73 images
- 1975 Mira Loma Revision - RC-1-16-1975 -- (RCFCWCD) 15 images
- 1975 Riverside Juvenile Hall - RC-3-5-1975 -- (RCFCWCD) 3 images
- 1975 University Wash - RC-6-17-1975 -- (RCFCWCD) 11 images
- 1975 University Wash Lake Evans - RC-1-9-1975 -- (RCFCWCD) 9 images
- 1975 Van Buren Sandpit - RC-2-4-1975 -- (RCFCWCD) 3 images
- 1976 Phoenix Avenue - RC-1-34-1976 -- (RCFCWCD) 31 images
- 1976 Rubidoux Disposal - RC-6-8-1976 -- (RCFCWCD) 3 images
- 1976 Spruce Street Storm Drain - RC-1-26-1976 -- (RCFCWCD) 26 images
- 1976 Spruce Street Storm Drain - RC-27-35-1976 -- (RCFCWCD) 9 images
- 1977 County Hospital - RC-1-4-1977 -- (RCFCWCD) 4 images
- 1977 Highgrove Disposal - RC-Highgrove-1-3-1977 -- (RCFCWCD) 3 images
- 1977 La Sierra - RC-1-49-1977 -- (RCFCWCD) 47 images
- 1977 Mira Loma Low Obliques - RC-1-2-1977 -- (RCFCWCD) 2 images
- 1977 Phoenix Avenue Storm Drain - RC-1-23-1977 -- (RCFCWCD) 23 images
- 1977 RCFC Office Low Obliques - RC-2-4-1977 -- (RCFCWCD) 3 images
- 1977 Rubidoux Disposal - RC-Rubidoux-1-3-1977 -- (RCFCWCD) 3 images
- 1977 Thornton Channel - RC-1-13-1977 -- (RCFCWCD) 13 images
- 1978 Alessandro Dam - RC-1-4-1978 -- (RCFCWCD) 4 images
- 1978 Arlington Channel - RC-1-5-1978 -- (RCFCWCD) 5 images
- 1978 Arlington Channel - RC-1-62-1978 -- (RCFCWCD) 62 images
- 1978 Box Springs Dam - RC-Box-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Harrison Dam - RC-3-5-1978 -- (RCFCWCD) 3 images
- 1978 High Grove - RC-High-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Poorman Dam - RC-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Prenda Dam - RC-Prenda-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Rubidoux Disposal - RC-Rubidoux-1-3-1978 -- (RCFCWCD) 3 images
- 1978 Sycamore / Arroyo Canyon - RC-1-6-1978 -- (RCFCWCD) 6 images
- 1978 Thornton Channel - RC-Thornton-1-10-1978 -- (RCFCWCD) 10 images
- 1978 Woodcrest Dam/Mabey Canyon - RC-1-2-1978 -- (RCFCWCD) 2 images
- 1978 Woodcrest Dam/Mabey Canyon - RC-Woodcrest-1-2-1978 -- (RCFCWCD) 2 images
- 1979 Alessandro Dam - RC-Alessandro-1-5-1979 -- (RCFCWCD) 5 images
- 1979 Box Springs Dam - RC-Box-1-3-1979 -- (RCFCWCD) 3 images
- 1979 Coachella Sanitary Landfill - RC-1-1-7-1979 -- (RCFCWCD) 5 images
- 1979 Day Creek Extention - RC-11-22-1979 -- (RCFCWCD) 12 images
- 1979 Harrison Dam - RC-Harrison-1-3-1979 -- (RCFCWCD) 3 images
- 1979 Highgrove - RC-Highgrove-2-3-5-1979 -- (RCFCWCD) 3 images
- 1979 Jamestown Drain - RC-1-12-1979 -- (RCFCWCD) 12 images
- 1979 Jamestown Drain - RC-1-14-1979 -- (RCFCWCD) 14 images
- 1979 Jamestown Drain - RC-Jameston-1-14-1979 -- (RCFCWCD) 14 images
- 1979 Mary Street Dam - RC-1-19-1979 -- (RCFCWCD) 19 images
- 1979 Monroe Channel - RC-Monroe-1-4-1979 -- (RCFCWCD) 4 images
- 1979 Monroe Railroad Lateral - RC-1-51-1979 -- (RCFCWCD) 51 images
- 1979 Prenda Dam - RC-Prenda-1-3-1979 -- (RCFCWCD) 3 images
- 1979 Rubidoux - RC-3-5-1979 -- (RCFCWCD) 6 images
- 1979 Sunnyslope Channel - RC-Sunnyslope-1-4-1979 -- (RCFCWCD) 4 images
- 1979 Sycamore Dam - RC-Sycamore-Dam-1-7-1979 -- (RCFCWCD) 7 images
- 1979 University Wash -RC-University-1-2-1979 -- (RCFCWCD) 2 images
- 1979 Woodcrest Dam/Mabey Canyon - RC-1-2-1979 -- (RCFCWCD) 2 images
- 1979 Woodcrest Dam/Mabey Canyon -RC-1-5-1979 -- (RCFCWCD) 5 images
- 1979 Woodcrest Dam - RC-Woodcrest-1-5-1979 -- (RCFCWCD) 5 images
- 1980 Alessandro Dam - RC-1-4-1980 -- (RCFCWCD) 4 images
- 1980 Arizona Channel - RC-14-30-1980 -- (RCFCWCD) 17 images
- 1980 Day Creek - RC-1-26-1980 -- (RCFCWCD) 26 images
- 1980 Harrison Dam - RC-1-3-1980 -- (RCFCWCD) 3 images
- 1980 Highgrove - RC-1-10-1980 -- (RCFCWCD) 10 images
- 1980 Highgrove - RC-Highgrove-1-5-1980 -- (RCFCWCD) 5 images
- 1980 Highgrove - RC-Highgrove-1-3-1980 -- (RCFCWCD) 3 images
- 1980 Jamestown Drain - RC-Jamestown-1-1980 -- (RCFCWCD) 1 images
- 1980 La Sierra Avenue Flood Damage - RC-1-7-1980 -- (RCFCWCD) 7 images
- 1980 Prenda Dam - RC-Prenda-1-3-1980 -- (RCFCWCD) 3 images
- 1980 Riverside Oblique - RC-Riverside-1-2-1980 -- (RCFCWCD) 2 images
- 1980 Sycamore Dam - RC-1-7-1980 -- (RCFCWCD) 7 images
- 1980 Woodcrest - RC-1-52-1980 -- (RCFCWCD) 31 images
- 1980 Woodcrest Dam - RC-1-5-1980 -- (RCFCWCD) 4 images
- 1980 Woodcrest Dam - RC-1-5-1980 -- (RCFCWCD) 5 images
- 1981 Arizona Channel - RC-Arizona-7-12-1981 -- (RCFCWCD) 6 images
- 1981 Mary Street Dam - RC-1-4-1981 -- (RCFCWCD) 4 images
- 1981 Mary Street Laterals - RC-1-30-1981 -- (RCFCWCD) 30 images
- 1981 South Riverside Ortho - RC-1-37-1981 -- (RCFCWCD) 37 images
- 1982 Alessandro Dam - RC-1-5-1982 -- (RCFCWCD) 5 images
- 1982 Harrison Dam - RC-Harrison-1-4-1982 -- (RCFCWCD) 4 images
- 1982 Mary Street Dam - RC-1-2-1982 -- (RCFCWCD) 2 images
- 1982 Mary Street Oblique - RC-1-1982 -- (RCFCWCD) 1 images
- 1982 Mary Street Dam - RC-3-4-1982 -- (RCFCWCD) 2 images
- 1982 Mary Street Dam - RC-Mary-1-4-1982 -- (RCFCWCD) 4 images
- 1982 Paramount Estates - RC-Paramount-1-2-1982-- (RCFCWCD) 2 images
- 1982 Prenda Dam - RC-Prenda-1-5-1982 -- (RCFCWCD) 5 images
- 1982 Riverside Area Obliques - RC-1-9-1982 -- (RCFCWCD) 9 images
- 1982 Rubidoux Sanitary Landfill - RC-1-3-1982 -- (RCFCWCD) 3 images
- 1982 Santa Ana River - RC-1-64-1982 -- (RCFCWCD) 64 images
- 1982 Woodcrest Dam - RC-1-4-1982 -- (RCFCWCD) 4 images
- 1983 Alessandro Basin - RC-Alessandro-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Box Springs Basin - RC-Box-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Harrison Basin - RC-Harrison-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Highgrove - RC-Highgrove2-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Highgrove Sanitary Landfill - RC-Highgrove-1-5-1983 -- (RCFCWCD) 5 images
- 1983 Jurupa - RC-1-21-1983 -- (RCFCWCD) 21 images
- 1983 Jurupa Basin - RC-Jurupa-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Jurupa Basin Outlet - RC-Jurupa-1-13-1983 -- (RCFCWCD) 13 images
- 1983 Jurupa - RC-Jurupa-2-4-1983 -- (RCFCWCD) 3 images
- 1983 Kansas Basin - RC-Kansas-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Marlborough Basin - RC-Marlborough-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Mary Street Basin - RC-Mary-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Monroe Basin - RC-Monroe-1-4-1983 -- (RCFCWCD) 4 images
- 1983 Prenda Basin - RC-Prenda-1-6-1983 -- (RCFCWCD) 6 images
- 1983 Rubidoux Basin - RC-Rubidoux2-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Rubidoux Sanitary Landfill - RC-Rubidoux-1-2-1983 -- (RCFCWCD) 2 images
- 1983 Sycamore Basin - RC-Sycamore2-1-6-1983 -- (RCFCWCD) 6 images
- 1983 Sycamore Dam - RC-Sycamore-1-6-1983 -- (RCFCWCD) 6 images
- 1983 Woodcrest Basin - RC-Woodcrest-1-5-1983 -- (RCFCWCD) 5 images
- 1984 Alessandro Basin - RC-Alessandro-1-5-1984 -- (RCFCWCD) 5 images
- 1984 Anza Basin - RC-Anza-1-2-1984 -- (RCFCWCD) 2 images
- 1984 Arlington Channel - RC-1-27-1984 -- (RCFCWCD) 26 images
- 1984 Box Springs Basin - RC-Box-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Columbia Basin - RC-Columbia-1-2-1984 -- (RCFCWCD) 2 images
- 1984 Harrison Basin - RC-Harrison-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Highgrove Sanitary Landfill - RC-Highgrove-1-14-1984 -- (RCFCWCD) 15 images
- 1984 Jurupa - Flight 135-84-1-1 -- (Imbiorski) 2 images
- 1984 Jurupa Basin - RC-Jurupa-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Kansas Basin - RC-Kansas-1-2-1984 -- (RCFCWCD) 2 images
- 1984 Marlborough Basin - RC-Marlborough-1-2-1984 -- (RCFCWCD) 2 images
- 1984 Mary Street Basin - RC-Mary-1-4-1984 -- (RCFCWCD) 4 images
- 1984 Monroe Basin - RC-Monroe-1-5-1984 -- (RCFCWCD) 5 images
- 1984 Pigeon Pass Road - RC-1-16-1984 -- (RCFCWCD) 16 images
- 1984 Prenda Basin - RC-Prenda-1-6-1984 -- (RCFCWCD) 6 images
- 1984 Riverside County Flood Control - RC-1-13-1984 -- (RCFCWCD) 13 images
- 1984 Rubidoux Basin - RC-Rubidoux-1-2-1984 -- (RCFCWCD) 2 images
- 1984 San Sevaine Channel - RC-Sevaine-1-9-1984 -- (RCFCWCD) 9 images
- 1984 San Sevaine Channel Extension - RC-1-12-1984 -- (RCFCWCD) 12 images
- 1984 San Sevaine Channel Extension - RC-Sevaine-1-22-1984 -- (RCFCWCD) 22 images
- 1984 South Riverside Channel - RC-South-1-3-19843 -- (RCFCWCD) 3 images
- 1984 Spring Street Storm Drain - RC-Spring-1-2-1984 -- (RCFCWCD) 2 images
- 1984 Sycamore Basin - RC-1-7-1984 -- (RCFCWCD) 7 images
- 1984 Woodcrest Basin -RC-Woodcrest-1-4-1984 -- (RCFCWCD) 4 images
- 1985 Alessandro Basin - RC-Alessandro-1-4-1985 -- (RCFCWCD) 4 images
- 1985 Anza Basin - RC-1-2-1985 -- (RCFCWCD) 2 images
- 1985 Box Springs Basin - RC-Box-1-4-1985 -- (RCFCWCD) 4 images
- 1985 City of Riverside - WO-224-85-1985 -- (Imbiorski) 9 images
- 1985 Columbia Basin - RC-Columbia-1-2-1985 -- (RCFCWCD) 2 images
- 1985 Glen Avon Hastings - RC-1-20-1985 -- (RCFCWCD) 20 images
- 1985 Harrison Basin - RC-Harrison-1-5-1985 -- (RCFCWCD) 5 images
- 1985 Highland Springs - RC-Highlands-1-18-1985 -- (RCFCWCD) 18 images
- 1985 Jurupa Basin - RC-Jurupa-1-3-1985 -- (RCFCWCD) 3 images
- 1985 Kansas Basin - RC-Kansas-1-2-1985 -- (RCFCWCD) 2 images
- 1985 Marlborough Basin - RC-Marlborough-1-2-1985 -- (RCFCWCD) 2 images
- 1985 Mary Street Basin - RC-Mary-1-4-1985 -- (RCFCWCD) 4 images
- 1985 Monroe Basin - RC-Monroe-1-4-1985 -- (RCFCWCD) 4 images
- 1985 Prenda Basin - RC-1-6-1985 -- (RCFCWCD) 6 images
- 1985 Riverside Bike Trail - RC-1-22-1985 -- (RCFCWCD) 21 images
- 1985 Riverside County District - RC-Riverside-1-5-1985 -- (RCFCWCD) 5 images
- 1985 Riverside Ortho - RC-1-91-1985 -- (RCFCWCD) 88 images
- 1985 Riverside Ortho -RC-1-3-1985 -- (RCFCWCD) 3 images
- 1985 Rubidoux Basin - RC-Rubidoux-1-2-1985 -- (RCFCWCD) 2 images
- 1985 South Lake Mathews - RC-1-27-1985 -- (RCFCWCD) 27 images
- 1985 Sycamore Basin - RC-1-7-1985 -- (RCFCWCD) 7 images
- 1985 Tilton Avenue Storm Drain - RC-1-5-1985 -- (RCFCWCD) 5 images
- 1985 Woodcrest Basin - RC-Woodcrest-1-4-1985 -- (RCFCWCD) 4 images
- 1986 Alessandro Basin - RC-1-5-1986 -- (RCFCWCD) 5 images
- 1986 Alessandro Basin - RC-1-4-1986 -- (RCFCWCD) 4 images
- 1986 Arlington Channel Stage 3 - RC-1-15-1986 -- (RCFCWCD) 15 images
- 1986 City of Riverside - WO-143-86-1986 -- Imbiorski 3 images
- 1986 City of Riverside - WO-265-86-1986 -- Imbiorski 28 images
- 1986 Harrison Basin - RC-Harrison-1-6-1986 -- (RCFCWCD) 5 images
- 1986 Jurupa Channel - RC-Jurupa-1-6-1986 -- (RCFCWCD) 6 images
- 1986 Limonite Study - RC-Limonite-1-4-1986 -- (RCFCWCD) 4 images
- 1986 Main Street Basin - RC-Main-1-5-1986 -- (RCFCWCD) 5 images
- 1986 Mission Boulevard Storm Drain - RC-Mission-1-4-1986 -- (RCFCWCD) 4 images
- 1986 Oak Street Basin - RC-Oak-1-4-1986 -- (RCFCWCD) 4 images
- 1986 Prenda Basin - RC-Prenda-1-6-1986 -- (RCFCWCD) 6 images
- 1986 San Sevaine Channel - RC-Sevaine-1-22-1986 -- (RCFCWCD) 22 images
- 1986 San Sevaine Channel - RC-1-24-1986 -- (RCFCWCD) 24 images
- 1986 Sycamore Basin - RC-Sycamore-1-7-1986 -- (RCFCWCD) 7 images
- 1986 Woodcrest Basin - RC-Woodcrest-1-9-1986 -- (RCFCWCD) 4 images
- 1987 Alessandro Basin - RC-Alessandro-1-5-1987 -- (RCFCWCD) 5 images
- 1987 Anza Basin - RC-Anza2-1-2-1987 -- (RCFCWCD) 2 images
- 1987 Bly Channel - RC-Bly-1-7-1987 -- (RCFCWCD) 7 images
- 1987 Box Springs Basin - RC-Box-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Columbia Basin - RC-Columbia-1-2-1987 -- (RCFCWCD) 2 images
- 1987 County Road Maintenance Yard - RC-1-1987 -- (RCFCWCD) 1 image
- 1987 Fairmont Park - RC-Fairmont-1-6-1986 -- (RCFCWCD) 6 images
- 1987 Harrison Basin - RC-Harrison-1-5-1987 -- (RCFCWCD) 5 images
- 1987 Jurupa Basin - RC-Jurupa-1-3-1987 -- (RCFCWCD) 3 images
- 1987 Kansas Basin - RC-Kansas-1-2-1987 -- (RCFCWCD) 2 images
- 1987 Marlborough Basin - RC-Marlborough-1-2-1987 -- (RCFCWCD) 2 images
- 1987 Mary Street Basin - RC-Mary-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Monroe Basin - RC-Monroe-1-5-1987 -- (RCFCWCD) 5 images
- 1987 Prenda Basin - RC-Prenda-1-6-1987 -- (RCFCWCD) 6 images
- 1987 Rubidoux Basin - RC-1-2-1987 -- (RCFCWCD) 2 images
- 1987 San Sevaine Channel - RC-Sevaine-1-24-1987 -- (RCFCWCD) 24 images
- 1987 Sycamore Basin - RC-1-7-1987 -- (RCFCWCD) 7 images
- 1987 West Riverside - RC-1-3-1987 -- (RCFCWCD) 3 images
- 1987 Woodcrest Basin - RC-Woodcrest-1-5-1987 -- (RCFCWCD) 5 images
- 1988 Alessandro Basin - RC-Alessandro-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Animal Shelter - RC-Animal-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Anza Basin - RC-Anza-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Arlington Drain Spot - RC-Arlington-1-1988 -- (RCFCWCD) 1 image
- 1988 Box Springs Basin - RC-Box-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Colton Sanitary Landfill - RC-Colton-1-6-1988 -- (RCFCWCD) 6 images
- 1988 Columbia Basin - RC-Columbia-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Day Creek - RC-Day-1-11-1988 -- (RCFCWCD) 11 images
- 1988 Harrison Basin - RC-Harrison-1-6-1988 -- (RCFCWCD) 6 images
- 1988 Highgrove Spot - RC-1-14-1988 -- (RCFCWCD) 14 images
- 1988 Highgrove Spot - RC-Highgrove-1-1988 -- (RCFCWCD) 1 images
- 1988 Highgrove - RC-Highgrove2-1-14-1988 -- (RCFCWCD) 10 images
- 1988 Jurupa Basin - RC-Jurupa-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Kansas Basin - RC-Kansas-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Marlborough Basin - RC-Marlborough-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Mary Street Basin - RC-Mary-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Monroe Basin - RC-Monroe-1-4-1988 -- (RCFCWCD) 4 images
- 1988 Prenda Basin - RC-Prenda-1-6-1988 -- (RCFCWCD) 6 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-I-1988 -- (RCFCWCD) 5 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-J-1988 -- (RCFCWCD) 9 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-K-1988 -- (RCFCWCD) 12 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-L-1988 -- (RCFCWCD) 10 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-M-1988 -- (RCFCWCD) 10 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-N-1988 -- (RCFCWCD) 9 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-O-1988 -- (RCFCWCD) 8 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-P-1988 -- (RCFCWCD) 6 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-Q-1988 -- (RCFCWCD) 6 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-R-1988 -- (RCFCWCD) 5 images
- 1988 Riverside / San Bernardino - RC-1-22-S-31-S-1988 -- (RCFCWCD) 3 images
- 1988 Rubidoux Basin - RC-Rubidoux-1-2-1988 -- (RCFCWCD) 2 images
- 1988 Sycamore Basin - RC-Sycamore-1-7-1988 -- (RCFCWCD) 7 images
- 1988 West Riverside - RC-Riverside-1-6-1988 -- (RCFCWCD) 6 images
- 1988 West Riverside Spot - RC-1-1988 -- (RCFCWCD) 1 image
- 1988 Woodcrest Basin - RC-Woodcrest-1-4-1988 -- (RCFCWCD) 4 images
- 1989 Anza Basin - RC-Anza-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Bellgrave-Etiwanda - RC-Bellgrave-1-1989 -- (RCFCWCD) 1 image
- 1989 Box Springs Basin - RC-Box-1-3-1989 -- (RCFCWCD) 3 images
- 1989 Columbia Basin - RC-Columbia-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Harrison Basin - RC-Harrison-1-5-1989 -- (RCFCWCD) 5 images
- 1989 Highgrove Spot - RC-Highgrove-1-1989 -- (RCFCWCD) 1 image
- 1989 Highgrove Street - RC-1-6-1989 -- (RCFCWCD) 6 images
- 1989 Jurupa Basin - RC-Jurupa-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Justice Center - RC-Justice-1-3-1989 -- (RCFCWCD) 3 images
- 1989 Justice Center - RC-Justice2-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Kansas Basin - RC-Kansas-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Limonite Avenue - RC-Limonite-1-10-1989 -- (RCFCWCD) 10 images
- 1989 Marlborough Basin - RC-Marlborough-1-2-1989 -- (RCFCWCD) 2 images
- 1989 Mary Street Basin - RC-Mary-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Mira Loma Ortho - RC-4-63-1989 -- (RCFCWCD) 59 images
- 1989 Monroe Basin - RC-Monroe-1-4-1989 -- (RCFCWCD) 4 images
- 1989 Prenda Basin - RC-Prenda-1-6-1989 -- (RCFCWCD) 6 images
- 1989 San Sevaine Channel - RC-1-21-1989 -- (RCFCWCD) 21 images
- 1989 Santa Ana River - RC-1-30-1989 -- (RCFCWCD) 29 images
- 1989 Southwest Riverside C-5 - RC-1-17-1989 -- (RCFCWCD) 17 images
- 1989 Sycamore Basin - RC-Sycamore-1-6-1989 -- (RCFCWCD) 6 images
- 1989 Woodcrest Basin - RC-Woodcrest-1-5-1989 -- (RCFCWCD) 5 images
- 1990 Alessandro Dam - RC-Alessandro-1-5-1990 -- (RCFCWCD) 5 images
- 1990 Anza Basin - RC-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Belltown Sanitary Landfill - RC-Belltown-2-1990 -- (RCFCWCD) 1 image
- 1990 Box Springs Dam - RC-Box-1-3-1990 -- (RCFCWCD) 3 images
- 1990 Columbia Basin - RC-Columbia-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Day Creek - RC-1-12-1990 -- (RCFCWCD) 12 images
- 1990 Harrison Dam - RC-Harrispn-1-5-1990 -- (RCFCWCD) 5 images
- 1990 Highgrove Sanitary Landfill - RC-1-14-1990 -- (RCFCWCD) 14 images
- 1990 Jurupa Basin - RC-Jurupa-1-3-1990 -- (RCFCWCD) 3 images
- 1990 Kansas Basin - RC-Kansas-1-2-1990 -- (RCFCWCD) 2 images
- 1990 Mary Street Basin - RC-Mary-1-4-1990 -- (RCFCWCD) 4 images
- 1990 Monroe Basin - RC-Monroe-1-4-1990 -- (RCFCWCD) 4 images
- 1990 Prenda Dam - RC-Prenda-1-6-1990 -- (RCFCWCD) 6 images
- 1990 Rubidoux Basin - RC-Rubidoux-1-2-1990 -- (RCFCWCD) 2 images
- 1990 San Sevaine - RC-1-37-1990 -- (RCFCWCD) 37 images
- 1990 Sycamore Dam - RC-Sycamore-1-6-1990 -- (RCFCWCD) 6 images
- 1990 Woodcrest Dam - RC-Woodcrest-1-4-1990 -- (RCFCWCD) 4 images
- 1991 Anza Basin - RC-Anza-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Arizona Channel - RC-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Box Springs - RC-Box-1-3-1991 -- (RCFCWCD) 3 images
- 1991 Canyon Crest - RC-1-10-1991 -- (RCFCWCD) 10 images
- 1991 Columbia Basin - RC-Columbia-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Day Creek Channel - RC-Day-1-7-1991 -- (RCFCWCD) 7 images
- 1991 Harrison Basin - RC-Harrison-1-5-1991 -- (RCFCWCD) 5 images
- 1991 Highgrove Sanitary Landfill - RC-1-14-1991 -- (RCFCWCD) 14 images
- 1991 Jurupa Basin - RC-Jurula-1-5-1991 -- (RCFCWCD) 4 images
- 1991 Kansas Basin - RC-Kansas-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Marlborough Basin - RC-Marlborough-1-2-1991 -- (RCFCWCD) 2 images
- 1991 Mary Street Basin - RC-Mary-1-4-1991 -- (RCFCWCD) 4 images
- 1991 Monroe Basin - RC-Monroe-1-5-1991 -- (RCFCWCD) 5 images
- 1991 Prenda Basin - RC-Prenda-1-6-1991 -- (RCFCWCD) 6 images
- 1991 Riverside / San Bernardino 1992 - RC-G-23-L-28-G-1991 -- (RCFCWCD) 5 images
- 1991 Riverside / San Bernardino 2007 - RC-G-23-L-28-H-1991 -- (RCFCWCD) 11 images
- 1991 Riverside / San Bernardino 2008 - RC-G-23-L-28-I-1991 -- (RCFCWCD) 11 images
- 1991 Riverside / San Bernardino 2019 - RC-G-23-L-28-J-1991 -- (RCFCWCD) 12 images
- 1991 Riverside / San Bernardino 2031 - RC-G-23-L-28-K-1991 -- (RCFCWCD) 13 images
- 1991 Riverside / San Bernardino 2044 - RC-G-23-L-28-L-1991 -- (RCFCWCD) 8 images
- 1991 Rubidoux Basin - RC-Rubidoux-1-2-1991 -- (RCFCWCD) 2 images
- 1991 San Sevaine Channel - RC-1-23-1991 -- (RCFCWCD) 23 images
- 1991 Santa Ana River - RC-1-28-1991 -- (RCFCWCD) 28 images
- 1991 Sherriffs Training Center - RC-1-6-1991 -- (RCFCWCD) 6 images
- 1991 Sycamore - RC-Sycamore-1-6-1991 -- (RCFCWCD) 6 images
- 1991 Woodcrest Basin - RC-Woodcrest-1-4-1991 -- (RCFCWCD) 4 images
- 1992 Alessandro Basin - RC-Alessandro-1-5-1992 -- (RCFCWCD) 5 images
- 1992 Arlington Channel Stage 4 - RC-Arlington-1-5-1992 -- (RCFCWCD) 5 images
- 1992 Arlington Channel Stage 5 - RC-Arlington-1-10-1992 -- (RCFCWCD) 10 images
- 1992 Box Springs Basin - RC-Box-1-3-1992 -- (RCFCWCD) 3 images
- 1992 Columbia Basin - RC-Columbia-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Harrison Basin - RC-Harrison-1-6-1992 -- (RCFCWCD) 6 images
- 1992 Highgrove Sanitary Landfill - RC-1-12-1992 -- (RCFCWCD) 12 images
- 1992 Highgrove Sanitary Landfill - RC-A-1992-A -- (RCFCWCD) 1 image
- 1992 Jurupa Basin - RC-Jurupa-1-3-1992 -- (RCFCWCD) 3 images
- 1992 Kansas BAsin - RC-Kansas-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Marlborough Basin - RC-Marlborough-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Mary Street Basin - RC-Mary-1-4-1992 -- (RCFCWCD) 4 images
- 1992 Monroe Basin - RC-Monroe-1-4-1992 -- (RCFCWCD) 4 images
- 1992 Pigeon Pass Basin - RC-Pigeon-1-8-1992 -- (RCFCWCD) 8 images
- 1992 Prenda Basin - RC-Prenda-1-6-1992 -- (RCFCWCD) 6 images
- 1992 Rubidoux Basin - RC-Rubidoux-1-2-1992 -- (RCFCWCD) 2 images
- 1992 Santa Ana River - RC-1-31-1992 -- (RCFCWCD) 31 images
- 1992 Sycamore Basin - RC-Sycamore-1-6-1992 -- (RCFCWCD) 6 images
- 1992 Woodcrest Basin - RC-Woodcrest-1-4-1992 -- (RCFCWCD) 4 images
- 1993 64th Street - RC-1-2-1993 -- (RCFCWCD) 2 images
- 1993 Alessandro Basin - RC-Alessandro-1-5-1993 -- (RCFCWCD) 5 images
- 1993 Belltown Storm Drain - RC-Belltown-1-4-1993 -- (RCFCWCD) 4 images
- 1993 Belltown Addition - RC-Belltown-5-8-1993 -- (RCFCWCD) 4 images
- 1993 Box Spring Basin - RC-Box-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Day Creek Channel Stage 6 - RC-Day-1-8-1993 -- (RCFCWCD) 6 images
- 1993 Highgrove Sanitary Landfill - RC-1-12-1993 -- (RCFCWCD) 12 images
- 1993 Highgrove Sanitary Landfill - RC-1-1993 -- (RCFCWCD) 1 image
- 1993 Jurupa Basin - RC-Jurupa-1-3-1993 -- (RCFCWCD) 3 images
- 1993 Mary Street Basin - RC-Mary-1-6-1993 -- (RCFCWCD) 6 images
- 1993 Monroe Basin - RC-Monroe-1-4-1993 -- (RCFCWCD) 4 images
- 1993 Prenda -RC-Prenda-1-6-1993 -- (RCFCWCD) 6 images
- 1993 Reche Canyon - RC-Reche-1-15-1993 -- (RCFCWCD) 15 images
- 1993 San Sevaine - RC-1-22-1993 -- (RCFCWCD) 22 images
- 1993 San Sevaine CH - RC-1-11-1993 -- (RCFCWCD) 12 images
- 1993 Santa Ana River - RC-Santa-1-33-1993 -- (RCFCWCD) 33 images
- 1993 Santa Ana River - RC-1-42-1993 -- (RCFCWCD) 41 images
- 1993 Santa Ana River - RC-1-73-1993 -- (RCFCWCD) 64 images
- 1993 Sycamore Basin - RC-Sycamore2-1-6-1993 -- (RCFCWCD) 6 images
- 1993 Sycamore Dam - RC-Sycamore-1-4-1993 -- (RCFCWCD) 4 images
- 1993 Woodcrest Basin - RC-Woodcrest-1-4-1993 -- (RCFCWCD) 4 images
- 1994 Alessandro Basin - RC-Alessandro-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Belltown Spot - RC-Belltown-A-1994 -- (RCFCWCD) 1 image
- 1994 Belltown Sanitary Landfill - RC-Belltown2-1-2-1994 -- (RCFCWCD) 2 images
- 1994 Box Springs - RC-Box-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Harrison Basin - RC-Harrison-1-5-1994 -- (RCFCWCD) 5 images
- 1994 Indian Canyon Dam Site - RC-Indian-1-2-1994 -- (RCFCWCD) 2 images
- 1994 Jurupa Line - RC-1-5-1994 -- (RCFCWCD) 5 images
- 1994 La Sierra Ortho - RC-1-48-1994 -- (RCFCWCD) 48 images
- 1994 Mary Street Basin - RC-Mary-1-5-1994 -- (RCFCWCD) 4 images
- 1994 Monroe Basin - RC-Monroe-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Prenda Basin - RC-1-7-1994 -- (RCFCWCD) 7 images
- 1994 San Sevaine Stage 7 - RC-San-1-2-1994 -- (RCFCWCD) 2 images
- 1994 San Sevaine Stage 7 - RC-Sevaine-1-6-1994 -- (RCFCWCD) 6 images
- 1994 Santa Ana River - RC-1-43-1994 -- (RCFCWCD) 43 images
- 1994 Sycamore Basin - RC-Sycamore-1-7-1994 -- (RCFCWCD) 7 images
- 1994 Woodcrest - RC-Woodcrest-1-4-1994 -- (RCFCWCD) 4 images
- 1995 Alessandro Dam - RC-Alessandro-1-5-1995 -- (RCFCWCD) 5 images
- 1995 Belltown Sanitary Landfill - RC-Belltown2-1-3-1995 -- (RCFCWCD) 2 images
- 1995 Belltown Sanitary Landfill Spot - RC-Belltown-A-1995 -- (RCFCWCD) 1 image
- 1995 Box Springs Dam - RC-Box-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Day Creek - RC-Day-1-8-1995 -- (RCFCWCD) 8 images
- 1995 Harrison Dam - RC-Harrison-1-5-1995 -- (RCFCWCD) 5 images
- 1995 Highgrove Sanitary Landfill - RC-Highgrove2-1-12-1995 -- (RCFCWCD) 12 images
- 1995 Highgrove Sanitary Landfill Spot -RC-Highgrove-A-1995 -- (RCFCWCD) 1 image
- 1995 Main Street Basin - RC-Main-1-5-1995 -- (RCFCWCD) 5 images
- 1995 Mary Street Dam - RC-Mary-1-4-1995 -- (RCFCWCD) 4 images
- 1995 Oak Street Basin - RC_Oak-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Prenda Dam - RC-Prenda-1-6-1995 -- (RCFCWCD) 6 images
- 1995 San Sevaine Channel - RC-1-17-1995 -- (RCFCWCD) 17 images
- 1995 Santa Ana River - RC-1-36-1995 -- (RCFCWCD) 36 images
- 1995 Sycamore Dam - RC-Sycamore-1-6-1995 -- (RCFCWCD) 6 images
- 1995 West Riverside Sanitary Landfill - RC-West3-1-4-1995 -- (RCFCWCD) 4 images
- 1995 West Riverside Sanitary Landfill Spot - RC-West2-A-1995 -- (RCFCWCD) 1 image
- 1995 Woodcrest Dam - RC-Woodcrest-1-4-1995 -- (RCFCWCD) 4 images
- 1996 64th Street Storm Drain - RC-64th-1-4-1996 -- (RCFCWCD) 4 images
- 1996 Alessandro Basin - RC-Alessandro-1-5-1996 -- (RCFCWCD) 5 images
- 1996 Belltown Sanitary Landfill - RC-Belltown-1-2-1996 -- (RCFCWCD) 2 images
- 1996 Box Springs Dam - RC-Box-1-3-1996 -- (RCFCWCD) 3 images
- 1996 Day Creek/Martin Street Storm Drain - RC-1-5-1996 -- (RCFCWCD) 5 images
- 1996 Declez Basin - RC-Declez-1-4-1996 -- (RCFCWCD) 4 images
- 1996 Fontana Lateral - RC-1-6-1996 -- (RCFCWCD) 6 images
- 1996 Harrison Basin - RC-Harrison-1-5-1996 -- (RCFCWCD) 5 images
- 1996 Highgrove Sanitary Landfill - RC-Highgrove2-1-13-1996 -- (RCFCWCD) 13 images
- 1996 Highgrove Sanitary Landfill Spot - RC-Highgrove-A-1996 -- (RCFCWCD) 1 image
- 1996 Mary Street Dam - RC-Mary-1-4-1996 -- (RCFCWCD) 4 images
- 1996 Monroe Basin - RC-Monroe-1-4-1996 -- (RCFCWCD) 4 images
- 1996 Prenda Dam - RC-Prenda-1-6-1996 -- (RCFCWCD) 6 images
- 1996 Riverside I - RC-1-29-1996 -- (RCFCWCD) 29 images
- 1996 Riverside II - RC-1-30-1996 -- (RCFCWCD) 30 images
- 1996 San Sevaine - RC-Sevaine-1-8-1996 -- (RCFCWCD) 8 images
- 1996 Santa Ana River -RC-1-31-1996 -- (RCFCWCD) 31 images
- 1996 Sycamore basin - RC-Sycamore-1-6-1996 -- (RCFCWCD) 6 images
- 1996 West Riverside Sanitary Landfill - RC-West-1-4-1996 -- (RCFCWCD) 4 images
- 1996 Woodcrest Dam - RC-Woodcrest-1-4-1996 -- (RCFCWCD) 4 images
- 1997 Alessandro Basin - RC-Alessandro-1-5-1997 -- (RCFCWCD) 5 images
- 1997 Box Springs Basin - RC-Box-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Day Creek Channel - RC-1-8-1997 -- (RCFCWCD) 8 images
- 1997 Harrison Basin - RC-Harrison-1-5-1997 -- (RCFCWCD) 5 images
- 1997 Highgrove Sanitary Landfill - RC-Highgrove2-1-1-2-6-1997 -- (RCFCWCD) 12 images
- 1997 Highgrove Sanitary Landfill Spot - RC-Highgrove-1-1997 -- (RCFCWCD) 1 image
- 1997 Mary Street Basin - RC-Mary-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Monroe Basin - RC-Monroe-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Prenda Basin - RC-Prenda-1-6-1997 -- (RCFCWCD) 6 images
- 1997 Rubidoux Channel Street - RC-Rubidoux-1-3-1997 -- (RCFCWCD) 3 images
- 1997 San Sevaine Channel - RC-1-35-1997 -- (RCFCWCD) 35 images
- 1997 Santa Ana River - RC-1-36-1997 -- (RCFCWCD) 37 images
- 1997 Southwest Riverside Line G - RC-SW-1-4-1997 -- (RCFCWCD) 4 images
- 1997 Sycamore Basin - RC-1-7-1997 -- (RCFCWCD) 7 images
- 1997 Woodcrest Basin - RC-Woodcrest-1-4-1997 -- (RCFCWCD) 4 images
- 1998 Day Creek Channel - RC-1-9-1998 -- (RCFCWCD) 9 images
- 1998 Day Creek Main Drain Pipe Lateral A - RC-Day-1-6-1998 -- (RCFCWCD) 6 images
- 1998 Harrison Basin - RC-Harrison-1-1-5-1998 -- (RCFCWCD) 5 images
- 1998 Highgrove Sanitary Landfill - RC-1-12-1998 -- (RCFCWCD) 12 images
- 1998 Highgrove Sanitary Landfill Spot - RC-Highgrove-A-1998 -- (RCFCWCD) 1 image
- 1998 Jurupa Basin - RC-Jurupa-1-4-1998 -- (RCFCWCD) 4 images
- 1998 Martin Street Storm Drain Lateral A-L4 - RC-Martin-1-6-1998 -- (RCFCWCD) 6 images
- 1998 Mary Street Dam - RC-Mary-1-1-4-1998 -- (RCFCWCD) 4 images
- 1998 Monroe Basin - RC-Monroe-1-4-1998 -- (RCFCWCD) 4 images
- 1998 Prenda Dam Basin - RC-Prenda-1-6-1998 -- (RCFCWCD) 6 images
- 1998 Riverside 3 Ortho - RC-1-19-1998 -- (RCFCWCD) 19 images
- 1998 Rubidoux 30th Street Storm Drain - RC-Rubidoux-1-10-1998 -- (RCFCWCD) 10 images
- 1998 Santa Ana River - RC-1-35-1998 -- (RCFCWCD) 35 images
- 1998 Sycamore Basin - RC-Sycamore-1-6-1998 -- (RCFCWCD) 6 images
- 1998 Woodcrest Dam - RC-Woodcrest-1-4-1998 -- (RCFCWCD) 4 images
- 1999 Alessandro Basin - RC-Alessandro-1-5-1999 -- (RCFCWCD) 5 images
- 1999 Box Springs Basin - RC-Box-1-3-1999 -- (RCFCWCD) 3 images
- 1999 Day Creek Channel - RC-Day-1-8-1999 -- (RCFCWCD) 8 images
- 1999 De Clez Basin - RC-1-4-1999 -- (RCFCWCD) 4 images
- 1999 Fontana Lateral - RC-Fontana-1-6-1999 -- (RCFCWCD) 6 images
- 1999 Harrison Basin - RC-Harrison-1-5-1999 -- (RCFCWCD) 5 images
- 1999 Jurupa Basin - RC-Jurupa-1-5-1999 -- (RCFCWCD) 5 images
- 1999 Mary Street Basin - RC-Mary-1-4-1999 -- (RCFCWCD) 4 images
- 1999 Monroe Basin - RC-Monroe-1-4-1999 -- (RCFCWCD) 4 images
- 1999 Prenda Basin - RC-Prenda-1-6-1999 -- (RCFCWCD) 6 images
- 1999 San Sevaine Channel - RC-1-22-1999 -- (RCFCWCD) 22 images
- 1999 San Sevaine Channel - RC-1-37-1999 -- (RCFCWCD) 37 images
- 1999 Santa Ana River - RC-1-32-1999 -- (RCFCWCD) 32 images
- 1999 Sycamore Basin - RC-Sycamore-1-6-1999 -- (RCFCWCD) 6 images
- 1999 Woodcrest Basin - RC-Woodcrest-1-4-1999 -- (RCFCWCD) 4 images
- 2000 Alessandro Dam - RC-Alessandro-1-5-2000 -- (RCFCWCD) 5 images
- 2000 Box Springs - RC-Box-1-3-2000 -- (RCFCWCD) 3 images
- 2000 Day Creek Channel - RC-Day-1-8-2000 -- (RCFCWCD) 8 images
- 2000 De Clez Basin - RC-DeClez-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Eastvale Man Drain Pipe - RC-Eastvale-1-18-2000 -- (RCFCWCD) 18 images
- 2000 Fontana Lateral - RC-Fontana-1-6-2000 -- (RCFCWCD) 6 images
- 2000 Harrison Basin - RC-Harrison-1-5-2000 -- (RCFCWCD) 5 images
- 2000 Highgrove Sanitary Landfill - RC-Highgrove2-1-12-2000 -- (RCFCWCD) 12 images
- 2000 Highgrove Sanitary Landfill Spot - RC-Highgrove-A-2000 -- (RCFCWCD) 1 images
- 2000 Jurupa Basin - RC-Jurupa-1-5-2000 -- (RCFCWCD) 5 images
- 2000 Mary Street Dam - RC-Mary-1-4-2000 -- (RCFCWCD) 4 images
- 2000 Monroe Basin - RC-Monroe-1-5-2000 -- (RCFCWCD) 5 images
- 2000 Prenda Dam - RC-Prenda-1-6-2000 -- (RCFCWCD) 6 images
- 2000 San Sevaine Channel - RC-Sevaine-1-37-2000 -- (RCFCWCD) 37 images
- 2000 Santa Ana River - RC-1-33-2000 -- (RCFCWCD) 33 images
- 2000 Sycamore - RC-Sycamore-1-7-2000 -- (RCFCWCD) 7 images
- 2000 Woodcrest Basin - RC-Woodcrest-1-4-2000 -- (RCFCWCD) 4 images
- 2001 Alessandro Basin - RC-Alessandro-RC-1-5-2001 -- (RCFCWCD) 5 images
- 2001 Box Springs Basin - RC-Box-1-3-2001 -- (RCFCWCD) 3 images
- 2001 Harrison Basin - RC-Harrison-1-5-2001 -- (RCFCWCD) 5 images
- 2001 Highgrove Sanitary Landfill - RC-1-12-2001 -- (RCFCWCD) 12 images
- 2001 Jurupa Basin - RC-Jurupa-RC-1-5-2001 -- (RCFCWCD) 5 images
- 2001 La Sierra Main Drain Pipeline - RC-Sierra-1-7-2001 -- (RCFCWCD) 7 images
- 2001 Mary Street Basin - RC-Mary-1-5-2001 -- (RCFCWCD) 5 images
- 2001 Monroe Basin - RC-Monroe-1-4-2001 -- (RCFCWCD) 4 images
- 2001 Prenda Basin - RC-Prenda-1-6-2001 -- (RCFCWCD) 6 images
- 2001 Santa Ana River - RC-1-31-2001 -- (RCFCWCD) 31 images
- 2001 Sycamore Basin - RC-Sycamore-1-6-2001 -- (RCFCWCD) 6 images
- 2001 Woodcrest Basin - RC-Woodcrest-1-4-2001 -- (RCFCWCD) 4 images
- 2002 Alessandro Basin - RC-Alessandro-1-5-2002 -- (RCFCWCD) 5 images
- 2002 Day Creek Channel - RC-1-8-2002 -- (RCFCWCD) 8 images
- 2002 Jurupa Basin - RC-Jurupa-1-3-2002 -- (RCFCWCD) 3 images
- 2002 Mary Street Dam - RC-Mary-1-5-2002 -- (RCFCWCD) 5 images
- 2002 Monroe Basin - RC-Monroe-1-3-2002 -- (RCFCWCD) 3 images
- 2002 San Sevaine Channel - RC-1-38-2002 -- (RCFCWCD) 38 images
- 2002 San Sevaine Channel - RC-Sevaine-1-7-2002 -- (RCFCWCD) 7 images
- 2002 Santa Ana River - RC-1-25-2002 -- (RCFCWCD) 25 images
- 2002 Sycamore Dam - RC-1-6-2002 -- (RCFCWCD) 6 images
- 2002 Woodcrest Dam - RC-1-4-2002 -- (RCFCWCD) 4 images
- 2003 Alessandro Dam - RC-Alessandro-1-5-2003 -- (RCFCWCD) 5 images
- 2003 Anza Basin - RC-Anza-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Anza Basin - RC-Anza2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Belltown Market Street - RC-Belltown-1-14-2003 -- (RCFCWCD) 15 images
- 2003 Box Springs Basin - RC-Box-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Box Springs Dam - RC-Box-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Box Springs Dam - RC-Box2-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Box Springs Detention Basin - RC-Box2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Columbia Basin - RC-Columbia-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Day Creek Basin - RC-Day-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Day Creek Channel - RC-1-10-2003 -- (RCFCWCD) 10 images
- 2003 Day Creek Channel - RC-1-11-2003 -- (RCFCWCD) 11 images
- 2003 Day Creek Retention - RC-Day2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Harrison Dam - RC-Harrison2-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Harrison Dam - RC-Harrison-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Jurupa Basin - RC-Jurupa-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Marlborough Basin - RC-Marlborough2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Marlborough Basin - RC-Marlborough-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Mary Street Dam - RC-Mary-1-5-2003 -- (RCFCWCD) 5 images
- 2003 Mira Loma Beach Street Dam - RC-Mira-1-12-2003 -- (RCFCWCD) 12 images
- 2003 Monroe Basin - RC-Monroe-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Prenda Dam - RC-Prenda-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Riverside County Flood Control Headquarters - RC-RCFC-1-4-2003 -- (RCFCWCD) 4 images
- 2003 Rubidoux Detention Basin - RC-Rubidoux2-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Rubidoux Retention Basin - RC-Rubidoux-1-2-2003 -- (RCFCWCD) 2 images
- 2003 San Sevaine Channel - RC-Sevaine-1-7-2003 -- (RCFCWCD) 7 images
- 2003 Santa Ana River - RC-Santa-1-27-2003 -- (RCFCWCD) 27 images
- 2003 Sycamore Dam - RC-Sycamore-1-7-2003 -- (RCFCWCD) 7 images
- 2003 Woodcrest Dam - RC-Woodcrest-1-5-2003 -- (RCFCWCD) 5 images
- 2004 Alessandro Basin - RC-Alessandro-1-5-2004 -- (RCFCWCD) 5 images
- 2004 Anza Basin - RC-Anza-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Belltown Market Street Channel - RC-Belltown-13-19-2004 -- (RCFCWCD) 7 images
- 2004 Box Springs Dam - RC-Box-1-4-2004 -- (RCFCWCD) 4 images
- 2004 Box Springs Detention Basin - RC-Box2-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Columbia Basin - RC-Columbia-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Day Creek Channel - RC-1-10A-2004 -- (RCFCWCD) 12 images
- 2004 Day Creek Channel Stage 6 - RC-Stage-1-10-2004 -- (RCFCWCD) 10 images
- 2004 Day Creek Detention Basin - RC-Day-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Harrison Dam - RC-Harrison-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Jurupa Basin - RC-Jurupa-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Marlborough Basin - RC-Marlborough-1-2-2004 -- (RCFCWCD) 2 images
- 2004 Mary Street Dam - RC-Mary-1-5-2004 -- (RCFCWCD) 5 images
- 2004 Monroe Basin - RC-Monroe-1-3-2004 -- (RCFCWCD) 3 images
- 2004 Prenda Dam - RC-Prenda-1-4-2004 -- (RCFCWCD) 4 images
- 2004 Rubidoux Detention Basin - RC-Rubidoux-1-2-2004 -- (RCFCWCD) 2 images
- 2004 San Sevaine Channel - RC-Sevaine-1-7-2004 -- (RCFCWCD) 7 images
- 2004 Santa Ana River - RC-1-26-2004 -- (RCFCWCD) 26 images
- 2004 Sycamore Dam - RC-Sycamore-1-6-2004 -- (RCFCWCD) 6 images
- 2004 Woodcrest Dam - RC-Woodcrest-1-5-2004 -- (RCFCWCD) 1 image
- 2005 Alessandro Basin - RC-Alessandro7-1-5-2005 -- (RCFCWCD) 5 images
- 2005 Alessandro Basin - RC-Alessandro-1-5-2005 -- (RCFCWCD) 5 images
- 2005 Anza Basin - RC-Anza7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Belltown Sanitary Landfill - RC-Belltown-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Bolero Drive Storm Drain - RC-Bolero-1-4-2005 -- (RCFCWCD) 4 images
- 2005 Box Springs Dam - RC-Box7-1-4-2005 -- (RCFCWCD) 4 images
- 2005 Box Springs Detention Basin - RC-Box8-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Columbia Basin - RC-Columbia7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Day Creek Channel - RC-Day8-1-10-2005 -- (RCFCWCD) 10 images
- 2005 Day Creek Detention Basin - RC-Day7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Harrison Basin - RC-Harrison7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Jurupa Basin - RC-Jurupa7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Marlborough Basin - RC-Marlborough7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Mary Street Dam - RC-Mary7-1-5-2005 -- (RCFCWCD) 5 images
- 2005 Monroe Basin - RC-Monroe7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Prenda Dam - RC-Prenda7-1-3-2005 -- (RCFCWCD) 3 images
- 2005 Riverside Townsite - RC-1-1-2-2-2005 -- (RCFCWCD) 4 images
- 2005 Riverside Townsite - RC-Spot-2005 -- (RCFCWCD) 1 image
- 2005 Rubidoux Retention Basin - RC-Rubidoux7-1-2-2005 -- (RCFCWCD) 2 images
- 2005 San Jacinto Stage 4 - RC-1-104-2005 -- (RCFCWCD) 104 images
- 2005 San Sevaine Channel - RC-Sevaine7-1-7-2005 -- (RCFCWCD) 7 images
- 2005 Santa Ana River - RC-Santa-1-26-2005 -- (RCFCWCD) 26 images
- 2005 Santa Ana River - -RC-Santa7-1-26-2005 -- (RCFCWCD) 26 images
- 2005 Santa Ana River Rubidoux - RC-1-20-2005 -- (RCFCWCD) 20 images
- 2005 Santa Ana River Eastvale - RC-Eastvale-1-15-2005 -- (RCFCWCD) 15 images
- 2005 Sycamore Dam - RC-Sycamore7-1-6-2005 -- (RCFCWCD) 6 images
- 2005 Woodcrest Dam - RC-Woodcrest7-1-4-2005 -- (RCFCWCD) 4 images
- 2006 Anza Basin - RC-Anza-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Box Springs Dam - RC-Box-1-4-2006 -- (RCFCWCD) 4 images
- 2006 Box Springs Detention Basin - RC-Box2-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Columbia Basin - RC-Columbia6-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Day Creek Basin - RC-Day-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Day Creek Channel - RC-Day2-1-10-2006 -- (RCFCWCD) 10 images
- 2006 Harrison Dam - RC-Harrison-1-3-2006 -- (RCFCWCD) 3 images
- 2006 I-10 West Ortho - RC-1-33-2006 -- (RCFCWCD) 33 images
- 2006 Jurupa Basin - RC-Jurupa-1-3-2006 -- (RCFCWCD) 3 images
- 2006 La Sierra Ortho - RC-1-64-2006 -- (RCFCWCD) 64 images
- 2006 Marlborough Basin - RC-Marlborough-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Mary Street Dam - RC-Mary2-1-5-2006 -- (RCFCWCD) 5 images
- 2006 Mary Street Dam to Alessandro - RC-1-12-2006 -- (RCFCWCD) 12 images
- 2006 Monroe Basin - RC-Monroe-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Prenda Dam - RC-Prenda-1-3-2006 -- (RCFCWCD) 3 images
- 2006 Rubidoux Basin - RC-Rubidoux-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Santa Ana River - RC-Santa-1-25-2006 -- (RCFCWCD) 25 images
- 2006 Southwest Riverside Main Drain Pipe L-C Stage 5 - RC-Southwest-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Southwest Riverside Main Drain Pipe L-C Stage 5 - RC-Southwest2-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Sycamore Dam - RC-Sycamore-1-7-2006 -- (RCFCWCD) 7 images
- 2006 Woodcrest Dam - RC-Woodcrest-1-4-2006 -- (RCFCWCD) 4 images
- 2007 Alessandro Dam - RC-1-8-2007 -- (RCFCWCD) 8 images
- 2007 Alessandro Dam - RC-Alessandro-1-7-2007 -- (RCFCWCD) 7 images
- 2007 Anza Basin - RC-Anza-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Box Springs Dam - RC-Box2-1-4-2007 -- (RCFCWCD) 4 images
- 2007 Box Springs Detention Basin - RC-Box-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Columbia - RC-Columbia-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Day Creek Channel - RC-1-10-2007 -- (RCFCWCD) 10 images
- 2007 Day Creek Detention Basin - RC-Day -1-2-2007 -- (RCFCWCD) 2 images
- 2007 Harrison Dam - RC-Harrison-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Jurupa Basin - RC-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Marlborough Basin - RC-Marlborough-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Mary Street Dam - RC-Mary-1-5-2007 -- (RCFCWCD) 4 images
- 2007 Monroe Basin - RC-Monroe-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Prenda Dam - RC-Prenda-1-3-2007 -- (RCFCWCD) 3 images
- 2007 Riverside Ortho - RC-1-30-2007 -- (RCFCWCD) 30 images
- 2007 Rubidoux Detention Basin - RC-Rubidoux-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Santa Ana River - RC-Santa-1-25-2007 -- (RCFCWCD) 25 images
- 2007 Southwest Riverside Main Drain Pipe L-C Stage 5 - RC-MDP-1-2-2007 -- (RCFCWCD) 2 images
- 2007 Sycamore - RC-Sycamore-1-7-2007 -- (RCFCWCD) 7 images
- 2007 Woodcrest Dam - RC-Woodcrest-1-4-2007 -- (RCFCWCD) 4 images
- 2008 Alessandro Dam - RC-Alessandro7-1-7-2008 -- (RCFCWCD) 7 images
- 2008 Anza Basin - RC-Anza-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Box Springs Dam - RC-Box7-1-4-2008 -- (RCFCWCD) 4 images
- 2008 Box Springs Detention Basin - RC-Box8-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Columbia Basin - RC-Columbia7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Day Creek Detention Basin - RC-Day7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Day Creek Channel - RC-Day8-1-10-2008 -- (RCFCWCD) 10 images
- 2008 Harrison Dam - RC-Harrison7-1-3-2008 -- (RCFCWCD) 2 images
- 2008 Jurupa Basin - RC-Jurupa7-1-3-2008 -- (RCFCWCD) 3 images
- 2008 Marlborough Basin - RC-Marlborough7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Mary Street Dam - RC-Mary7-1-5-2008 -- (RCFCWCD) 5 images
- 2008 Monroe Basin - RC-Monroe-1-3-2008 -- (RCFCWCD) 3 images
- 2008 Prenda Dam - RC-Prenda7-1-3-2008 -- (RCFCWCD) 3 images
- 2008 Riverside County Flood Control & Water Conservation District Facility - RC-1-5-2008 -- (RCFCWCD) 5 images
- 2008 Riverside Ortho - RC-1-1-9-9-2008 -- (RCFCWCD) 103 images
- 2008 Rubidoux Retention Basin - RC-Rubidoux7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Santa Ana River - RC-Santa7-1-25-2008 -- (RCFCWCD) 25 images
- 2008 Southwest Riverside Main Drain Pipe - RC-Riverside7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Sycamore Dam - RC-Sycamore7-1-6-2008 -- (RCFCWCD) 6 images
- 2008 Woodcrest Dam - RC-Woodcrest7-1-4-2008 -- (RCFCWCD) 4 images
- 2009 Alessandro Dam - RC-Alessandro-1-7-2009 -- (RCFCWCD) 7 images
- 2009 Alessandro Dam Access - RC-Alessandro-1-19-2009 -- (RCFCWCD) 19 images
- 2009 Box Springs Dam - RC-Box-1-4-2009 -- (RCFCWCD) 4 images
- 2009 Day Creek Channel - RC-Day7-1-10-2009 -- (RCFCWCD) 10 images
- 2009 Day Creek Frank Avenue Storm Drain - RC-Day7-1-6-2009 -- (RCFCWCD) 6 images
- 2009 Day Creek Retention Basin - RC-Day8-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Harrison Dam - RC-Harrison-1-3-2009 -- (RCFCWCD) 3 images
- 9 Highgrove Channel Repair Work - RC-Highgrove7-1-8-2009 -- (RCFCWCD) 8 images
- 2009 Mary Street Dam - RC-Mary-1-5-2009 -- (RCFCWCD) 5 images
- 2009 Monroe Basin - RC-Monroe-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Prenda Dam - RC-Prenda7-1-3-2009 -- (RCFCWCD) 3 images
- 2009 Rubidoux retention Basin - RC-Rubidoux7-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Santa Ana River Digital Index - RC-1-14-2009 -- (RCFCWCD) 14 images
- 2009 Sycamore Dam - RC-Sycamore-1-6-2009 -- (RCFCWCD) 6 images
- 2009 Woodcrest Dam - RC-Woodcrest-1-4-2009 -- (RCFCWCD) 4 images
- 2010 Alessandro Dam - RC-Alessandro-1-3-2010 -- (RCFCWCD) 3 images
- 2010 Harrison Dam - RC-Harrison-1-3-2010 -- (RCFCWCD) 3 images
- 2010 Mary Street Dam - RC-Mary-1-3-2010 -- (RCFCWCD) 3 images
- 2010 Monroe Main Drain Pipe - RC-1-8-2010 -- (RCFCWCD) 8 images
- 2010 Santa Ana River - RC-1-39-2010 -- (RCFCWCD) 39 images
- 2010 Sycamore Dam - RC-1-7-2010 -- (RCFCWCD) 7 images
- 2010 Sycamore Dam - RC-Sycamore-1-3-2010 -- (RCFCWCD) 3 images
- 2011 Alessandro Dam - RC-Alessandro-1-7-2011 -- (RCFCWCD) 7 images
- 2011 Anza Basin - RC-Anza-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Box Springs Dam - RC-Box-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Box Springs Detention Basin - RC-Box-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Columbia basin - RC-Columbia-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Day Creek Detention - RC-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Harrison Dam - RC-Harrison-1-3-2011-- (RCFCWCD) 3 images
- 2011 Jurupa Basin - RC-Jurupa-1-3-2011-- (RCFCWCD) 3 images
- 2011 Marlborough Basin - RC-Marlborough-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Mary Street Dam - RC-1-5-2011-- (RCFCWCD) 5 images
- 2011 Monroe Basin - RC-1-4-2011 -- (RCFCWCD) 4 images
- 2011 Prenda Dam- RC-Prenda-1-3-2011-- (RCFCWCD) 3 images
- 2011 Reche Canyon - RC-1-29-2011-- (RCFCWCD) 29 images
- 2011 Rubidoux Detention Basin - RC-Rubidoux-1-2-2011-- (RCFCWCD) 2 images
- 2011 Santa Ana River - RC-1-16-2011 -- (RCFCWCD) 16 images
- 2011 Southwest Riverside Main Drain Pipeline - RC-1-9-2011 -- (RCFCWCD) 9 images
- 2011 Southwest Riverside Main Drain Pipeline Line C - RC-Line-1-2-2011 -- (RCFCWCD) 2 images
- 2011 University West Channel, Stage 3 - RC-University-1-3-2011 -- (RCFCWCD) 3 images
- 2011 Woodcrest Dam - RC-Woodcrest-1-4-2011 -- (RCFCWCD) 4 images
- 2012 Alessandro Dam - RC-Alessandro-1-7-2012 -- (RCFCWCD) 7 images
- 2012 Anza Basin - RC-Anza-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Box Springs Dam - RC-Box-1-4-2012 -- (RCFCWCD) 4 images
- 2012 Box Springs Detention Basin - RC-Box2-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Columbia Basin - RC-Columbia-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Day Creek Channel - RC-1-10-2012 -- (RCFCWCD) 10 images
- 2012 Day Creek Detention Basin - RC-Day2-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Harrison Dam - RC_Harrison-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Jurupa Basin - RC-Jurupa-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Marlborough Basin- RC-Marlborough-1-2-2012-- (RCFCWCD) 2 images
- 2012 Mary Street Dam - RC-Mary-1-5-2012 -- (RCFCWCD) 5 images
- 2012 Mockingbird Canyon Channel - RC-Mockingbird7-1-18-2012 -- (RCFCWCD) 18 images
- 2012 Monroe Basin - RC-Monroe-1-3-2012-- (RCFCWCD) 3 images
- 2012 Prenda Dam - RC-Prenda-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Riverside County Flood Control - RC-1-2012 -- (RCFCWCD) 1 image
- 2012 Rubidoux Detention Basin - RC-Rubidoux-1-2-2012 -- (RCFCWCD) 2 images
- 2012 San Sevaine Channel - RC-San9-1-3-2012 -- (RCFCWCD) 3 images
- 2012 Santa Ana River - RC-1-25-2012-- (RCFCWCD) 25 images
- 2012 Southwest River Main Drain Pipeline Line L, Stage 5 Basin - SW-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Sycamore Dam - RC-Sycamore-1-6-2012-- (RCFCWCD) 6 images
- 2012 Woodcrest Dam - RC-Woodcrest-1-4-2012 -- (RCFCWCD) 4 images
- 2013 Alessandro Basin Dam - RC-Alessandro-1-7-2013 -- (RCFCWCD) 7 images
- 2013 Anza Basin - RC-Anza-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Box Springs Dam - RC-Box-1-4-2013 -- (RCFCWCD) 4 images
- 2013 Box Springs Detention Basin - RC-Box2-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Columbia Basin - RC-Columbia-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Day Creek Channel - RC-1-10-2013 -- (RCFCWCD) 10 images
- 2013 Day Creek Detention Basin - RC-Day-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Harrison Dam - RC-Harrison-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Highgrove Sanitary Landfill - RC-Highgrove-1-12-2013 -- (RCFCWCD) 12 images
- 2013 Highgrove Sanitary Landfill Spot - RC-Highgrove-A-2013 -- (RCFCWCD) 1 image
- 2013 Jurupa Basin - RC-Jurupa-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Marlborough Basin - RC-Marlborough-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Mary Street Dam - RC-Mary-1-5-2013 -- (RCFCWCD) 5 images
- 2013 Monroe Basin - RC-Monroe-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Monroe Main Drain Pipeline Kine E, Stage 2 & 3 - RC-1-22-2013 -- (RCFCWCD) 22 image
- 2013 Prenda Dam- RC-Prenda-1-3-2013 -- (RCFCWCD) 3 images
- 2013 Rubidoux Basin- RC-Roubidoux-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Santa Ana River - RC-SAR-1-25-2013 -- (RCFCWCD) 25 images
- 2013 Southwest Riverside Main Drain Pipeline Line C - RC-SouthC-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Sycamore Dam Realignment - RC-Sycamore-1-6-2013 -- (RCFCWCD) 6 images
- 2013 Woodcrest Dam - RC-Wood-1-4-2013 -- (RCFCWCD) 4 image
- 2014 Riverside County Flood Control - RC-1-4-2014 -- (RCFCWCD) 4 images
- 2015 Alessandro Dam - RC-Alessandro-1-7-2015 -- (RCFCWCD) 7 images
- 2015 Anza Basin - RC-Anza-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Bellgrave Basin - RC-Bellgrave-1-3-2015 -- (RCFCWCD) 3 image
- 2015 Box Springs Dam - RC-Box-1-4-2015 -- (RCFCWCD) 4 images
- 2015 Box Springs Basin - RC-Box2-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Columbia Basin- RC-Columbia-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Day Creek Channel - RC-Day-1-10-2015 -- (RCFCWCD) 10 images
- 2015 Day Creek Detention Basin - RC-Day2-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Harrison Dam- RC-Harrison-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Jurupa Basin- RC-Jurupa-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Marlborough Basin - RC-Marlborough-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Mary Street Dam - RC-Mary-1-5-2015 -- (RCFCWCD) 5 images
- 2015 Monroe Basin - RC-Monroe-1-3-2015 -- (RCFCWCD) 3 images
- 2015 Rubidoux Detention Basin - RC-Rubidoux-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Santa Ana River - RC-SAR-1-25-2015 -- (RCFCWCD) 25 images
- 2015 Southwest Riverside Main Drain Pipeline Line C Stage 5 Basin - RC-Southwest-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Sycamore Dam - RC-Sycamore-1-6-2015 -- (RCFCWCD) 6 images
- 2015 Woodcrest Dam - RC-Woodcrest-1-4-2015 -- (RCFCWCD) 4 images
-
Riverside - County (17,042 images)
Riverside - County includes aerial photograph flights that cover the bulk of Riverside County rather than any of the designated zones. The flights start on the west side of Riverside County and lap back and forth in the north-south flight pattern.
- 1962 County of Riverside Flight - RC-3-270-1962 -- (RCFCWCD) 248 images
- 1962 County of Riverside Flight - RC-271-520-1962 -- (RCFCWCD) 244 images
- 1962 County of Riverside Flight - RC-521-567-1962 -- (RCFCWCD) 37 images
- 1974 County of Riverside Flight - RC-1-502-1974 - Part 1 -- (RCFCWCD) 252 images
- 1974 County of Riverside Flight - RC-1-502-1974 - Part 2 -- (RCFCWCD) 252 images
- 1974 County of Riverside Flight - RC-503-650-1974 - Part 3 -- (RCFCWCD) 252 images
- 1974 County of Riverside Flight - RC-651-1180-1974 - Part 4 -- (RCFCWCD) 252 images
- 1974 County of Riverside Flight - RC-651-1180-1974 - Part 5 -- (RCFCWCD) 172 images
- 1980 County of Riverside Flight - RC-1-1191-1980 - Part 1 -- (RCFCWCD) 249 images
- 1980 County of Riverside Flight - RC-1-1191-1980 - Part 2 -- (RCFCWCD) 246 images
- 1980 County of Riverside Flight - RC-1-1191-1980 - Part 3 -- (RCFCWCD) 249 images
- 1980 County of Riverside Flight - RC-1-1191-1980 - Part 4 -- (RCFCWCD) 247 images
- 1980 County of Riverside Flight - RC-1-1191-1980 - Part 5 -- (RCFCWCD) 191 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 1 -- (RCFCWCD) 250 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 2 -- (RCFCWCD) 249 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 3 -- (RCFCWCD) 250 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 4 -- (RCFCWCD) 248 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 5 -- (RCFCWCD) 250 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 6 -- (RCFCWCD) 250 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 7 -- (RCFCWCD) 250 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 8 -- (RCFCWCD) 250 images
- 1984 County of Riverside Flight - RC-1-1000-1984 - Part 9 -- (RCFCWCD) 145 images
- 1990 County of Riverside Flight - RC-1-85-1990-1 -- (RCFCWCD) 85 images
- 1990 County of Riverside Flight - RC-1-73-1990-2 -- (RCFCWCD) 73 image
- 1990 County of Riverside Flight - RC-1-81-1990-3 -- (RCFCWCD) 81 images
- 1990 County of Riverside Flight - RC-1-79-1990-4 -- (RCFCWCD) 79 images
- 1990 County of Riverside Flight - RC-1-93-1990-5 -- (RCFCWCD) 93 images
- 1990 County of Riverside Flight - RC-1-98-1990-6 -- (RCFCWCD) 98 images
- 1990 County of Riverside Flight - RC-1-90-1990-7 -- (RCFCWCD) 90 images
- 1990 County of Riverside Flight - RC-1-108-1990-8 -- (RCFCWCD) 108 images
- 1990 County of Riverside Flight - RC-1-98-1990-9 -- (RCFCWCD) 98 images
- 1990 County of Riverside Flight - RC-1-106-1990-10 -- (RCFCWCD) 106 images
- 1990 County of Riverside Flight - RC-1-92-1990-11 -- (RCFCWCD) 92 images
- 1990 County of Riverside Flight - RC-1-108-1990-12 -- (RCFCWCD) 108 images
- 1990 County of Riverside Flight - RC-1-96-1990-13 -- (RCFCWCD) 96 images
- 1990 County of Riverside Flight - RC-1-98-1990-14 -- (RCFCWCD) 98 images
- 1990 County of Riverside Flight - RC-1-101-1990-15 -- (RCFCWCD) 101 images
- 1990 County of Riverside Flight - RC-1-97-1990-16 -- (RCFCWCD) 97 images
- 1990 County of Riverside Flight - RC-1-101-1990-17 -- (RCFCWCD) 101 images
- 1990 County of Riverside Flight - RC-1-104-1990-18 -- (RCFCWCD) 104 images
- 1990 County of Riverside Flight - RC-1-89-1990-19 -- (RCFCWCD) 89 images
- 1990 County of Riverside Flight - RC-1-76-1990-20 -- (RCFCWCD) 76 images
- 1990 County of Riverside Flight - RC-1-76-1990-21 -- (RCFCWCD) 76 images
- 1990 County of Riverside Flight - RC-1-39-1990-22 -- (RCFCWCD) 39 images
- 1990 County of Riverside Flight - RC-1-102-1990-23 -- (RCFCWCD) 102 images
- 1990 County of Riverside Flight - RC-1-104-1990-24 -- (RCFCWCD) 104 images
- 1990 County of Riverside Flight - RC-1-92-1990-25 -- (RCFCWCD) 92 images
- 1990 County of Riverside Flight - RC-1-80-1990-26 -- (RCFCWCD) 80 images
- 1990 County of Riverside Flight - RC-1-85-1990-27 -- (RCFCWCD) 85 images
- 1990 County of Riverside Flight - RC-1-91-1990-28 -- (RCFCWCD) 92 images
- 1990 County of Riverside Flight - RC-1-81-1990-29 -- (RCFCWCD) 80 images
- 1990 County of Riverside Flight - RC-1-78-1990-30 -- (RCFCWCD) 78 images
- 1990 County of Riverside Flight - RC-1-59-1990-31 -- (RCFCWCD) 59 images
- 1990 County of Riverside Flight - RC-1-52-1990-32 -- (RCFCWCD) 52 images
- 1990 County of Riverside Flight - RC-1-53-1990-33 -- (RCFCWCD) 53 images
- 1990 County of Riverside Flight - RC-1-50-1990-34 -- (RCFCWCD) 50 images
- 1990 County of Riverside Flight - RC-1-50-1990-35 -- (RCFCWCD) 50 images
- 1990 County of Riverside Flight - RC-1-46-1990-36 -- (RCFCWCD) 51 images
- 1990 County of Riverside Flight - RC-1-49-1990-37 -- (RCFCWCD) 49 images
- 1990 County of Riverside Flight - RC-1-50-1990-38 -- (RCFCWCD) 50 images
- 1990 County of Riverside Flight - RC-1-50-1990-39 -- (RCFCWCD) 50 images
- 1990 County of Riverside Flight - RC-1-20-1990-40 -- (RCFCWCD) 20 images
- 1990 County of Riverside Flight - RC-1-19-1990-41 -- (RCFCWCD) 19 images
- 1990 County of Riverside Flight - RC-1-19-1990-42 -- (RCFCWCD) 19 images
- 1990 County of Riverside Flight - RC-1-18-1990-43 -- (RCFCWCD) 18 images
- 1990 County of Riverside Flight - RC-1-18-1990-44 -- (RCFCWCD) 18 images
- 1990 County of Riverside Flight - RC-1-20-1990-45 -- (RCFCWCD) 20 images
- 1990 County of Riverside Flight - RC-1-19-1990-46 -- (RCFCWCD) 19 images
- 1990 County of Riverside Flight - RC-1-17-1990-47 -- (RCFCWCD) 17 images
- 1990 County of Riverside Flight - RC-1-15-1990-48 -- (RCFCWCD) 15 images
- 1990 County of Riverside Flight - RC-1-16-1990-49 -- (RCFCWCD) 16 images
- 1990 County of Riverside Flight - RC-1-13-1990-50 -- (RCFCWCD) 13 images
- 1995 County of Riverside Flight - RC-1-1-69-1995-1 -- (RCFCWCD) 69 images
- 1995 County of Riverside Flight - RC-2-1-71-1995-2 -- (RCFCWCD) 69 images
- 1995 County of Riverside Flight - RC-3-1-74-1995-3 -- (RCFCWCD) 73 images
- 1995 County of Riverside Flight - RC-4-1-78-1995-4 -- (RCFCWCD) 78 images
- 1995 County of Riverside Flight - RC-5-1-81-1995-5 -- (RCFCWCD) 81 images
- 1995 County of Riverside Flight - RC-6-1-85-1995-6 -- (RCFCWCD) 85 images
- 1995 County of Riverside Flight - RC-7-1-88-1995-7 -- (RCFCWCD) 86 images
- 1995 County of Riverside Flight - RC-8-1-82-1995 -- (RCFCWCD) 82 images
- 1995 County of Riverside Flight - RC-9-1-87-1995 -- (RCFCWCD) 88 images
- 1995 County of Riverside Flight - RC-10-1-87-1995 -- (RCFCWCD) 87 images
- 1995 County of Riverside Flight - RC-11-1-85-1995 -- (RCFCWCD) 82 images
- 1995 County of Riverside Flight - RC-12-1-90-1995 -- (RCFCWCD) 90 images
- 1995 County of Riverside Flight - RC-13-1-91-1995 -- (RCFCWCD) 90 images
- 1995 County of Riverside Flight - RC-14-1-95-1995 -- (RCFCWCD) 95 images
- 1995 County of Riverside Flight - RC-15-1-92-1995 -- (RCFCWCD) 92 images
- 1995 County of Riverside Flight - RC-16-1-93-1995 -- (RCFCWCD) 90 images
- 1995 County of Riverside Flight - RC-17-1-94-1995 -- (RCFCWCD) 90 images
- 1995 County of Riverside Flight - RC-18-1-100-1995 -- (RCFCWCD) 100 images
- 1995 County of Riverside Flight - RC-19-1-81-1995 -- (RCFCWCD) 79 images
- 1995 County of Riverside Flight - RC-20-1-74-1995 -- (RCFCWCD) 72 images
- 1995 County of Riverside Flight - RC-21-1-76-1995 -- (RCFCWCD) 76 images
- 1995 County of Riverside Flight - RC-22-1-40-1995 -- (RCFCWCD) 40 images
- 1995 County of Riverside Flight - RC-23-1-86-1995 -- (RCFCWCD) 86 images
- 1995 County of Riverside Flight - RC-24-1-85-1995 -- (RCFCWCD) 85 images
- 1995 County of Riverside Flight - RC-25-1-84-1995 -- (RCFCWCD) 83 images
- 1995 County of Riverside Flight - RC-26-1-84-1995 -- (RCFCWCD) 83 images
- 1995 County of Riverside Flight - RC-27-1-84-1995 -- (RCFCWCD) 84 images
- 1995 County of Riverside Flight - RC-28-1-88-1995 -- (RCFCWCD) 88 images
- 1995 County of Riverside Flight - RC-29-1-65-1995 -- (RCFCWCD) 65 images
- 1995 County of Riverside Flight - RC-30-1-60-1995 -- (RCFCWCD) 60 images
- 1995 County of Riverside Flight - RC-31-1-58-1995 -- (RCFCWCD) 57 images
- 1995 County of Riverside Flight - RC-32-1-55-1995 -- (RCFCWCD) 56 images
- 1995 County of Riverside Flight - RC-33-1-58-1995 -- (RCFCWCD) 56 images
- 1995 County of Riverside Flight - RC-34-1-49-1995 -- (RCFCWCD) 48 images
- 1995 County of Riverside Flight - RC-35-1-52-1995 -- (RCFCWCD) 52 images
- 1995 County of Riverside Flight - RC-36-1-57-1995 -- (RCFCWCD) 57 images
- 1995 County of Riverside Flight - RC-37-1-52-1995 -- (RCFCWCD) 52 images
- 1995 County of Riverside Flight - RC-38-1-48-1995 -- (RCFCWCD) 48 images
- 1995 County of Riverside Flight - RC-39-1-49-1995 -- (RCFCWCD) 49 images
- 1995 County of Riverside Flight - RC-40-1-17-1995 -- (RCFCWCD) 17 images
- 1995 County of Riverside Flight - RC-41-1-19-1995 -- (RCFCWCD) 19 images
- 1995 County of Riverside Flight - RC-42-1-20-1995 -- (RCFCWCD) 20 images
- 1995 County of Riverside Flight - RC-43-1-22-1995 -- (RCFCWCD) 22 images
- 1995 County of Riverside Flight - RC-44-1-20-1995 -- (RCFCWCD) 20 images
- 1995 County of Riverside Flight - RC-45-1-17-1995 -- (RCFCWCD) 17 images
- 1995 County of Riverside Flight - RC-46-1-19-1995 -- (RCFCWCD) 19 images
- 1995 County of Riverside Flight - RC-47-1-18-1995 -- (RCFCWCD) 18 images
- 1995 County of Riverside Flight - RC-48-1-19-1995 -- (RCFCWCD) 20 images
- 1995 County of Riverside Flight - RC-49-1-19-1995 -- (RCFCWCD) 19 images
- 1995 County of Riverside Flight - RC-50-1-17-1995 -- (RCFCWCD) 17 images
- 1995 County of Riverside Flight - RC-51-1-14-1995 -- (RCFCWCD) 14 images
- 1995 County of Riverside Flight - RC-52-1-14-1995 -- (RCFCWCD) 14 images
- 2000 County of Riverside Flight - RC-1-1-73-2000 -- (RCFCWCD) 73 images
- 2000 County of Riverside Flight - RC-2-1-77-2000 -- (RCFCWCD) 77 images
- 2000 County of Riverside Flight - RC-3-1-79-2000 -- (RCFCWCD) 79 images
- 2000 County of Riverside Flight - RC-4-1-80-2000 -- (RCFCWCD) 79 images
- 2000 County of Riverside Flight - RC-5-1-84-2000 -- (RCFCWCD) 84 images
- 2000 County of Riverside Flight - RC-6-1-92-2000 -- (RCFCWCD) 92 images
- 2000 County of Riverside Flight - RC-7-1-93-2000 -- (RCFCWCD) 90 images
- 2000 County of Riverside Flight - RC-8-1-90-2000 -- (RCFCWCD) 89 images
- 2000 County of Riverside Flight - RC-9-1-90-2000 -- (RCFCWCD) 90 images
- 2000 County of Riverside Flight - RC-10-1-90-2000 -- (RCFCWCD) 90 images
- 2000 County of Riverside Flight - RC-11-1-92-2000 -- (RCFCWCD) 92 images
- 2000 County of Riverside Flight - RC-12-1-93-2000 -- (RCFCWCD) 93 images
- 2000 County of Riverside Flight - RC-13-1-93-2000 -- (RCFCWCD) 93 images
- 2000 County of Riverside Flight - RC-14-1-94-2000 -- (RCFCWCD) 94 images
- 2000 County of Riverside Flight - RC-15-1-97-2000 -- (RCFCWCD) 97 images
- 2000 County of Riverside Flight - RC-16-1-93-2000 -- (RCFCWCD) 94 images
- 2000 County of Riverside Flight - RC-17-1-100-2000 -- (RCFCWCD) 100 images
- 2000 County of Riverside Flight - RC-18-1-104-2000 -- (RCFCWCD) 104 images
- 2000 County of Riverside Flight - RC-19-1-84-2000 -- (RCFCWCD) 85 images
- 2000 County of Riverside Flight - RC-20-1-78-2000 -- (RCFCWCD) 78 images
- 2000 County of Riverside Flight - RC-21-1-74-2000 -- (RCFCWCD) 74 images
- 2000 Inland Empire Photo Atlas - RC-544-979-2000 -- (RCFCWCD) 100 images
- 2005 County of Riverside Flight - RC-1-1-78-2005 -- (RCFCWCD) 78 images
- 2005 County of Riverside Flight - RC-2-1-79-2005 -- (RCFCWCD) 79 images
- 2005 County of Riverside Flight - RC-3-1-88-2005 -- (RCFCWCD) 87 images
- 2005 County of Riverside Flight - RC-4-1-85-2005 -- (RCFCWCD) 85 images
- 2005 County of Riverside Flight - RC-5-1-83-2005 -- (RCFCWCD) 81 images
- 2005 County of Riverside Flight - RC-6-1-91-2005 -- (RCFCWCD) 82 images
- 2005 County of Riverside Flight - RC-7-1-88-2005 -- (RCFCWCD) 86 images
- 2005 County of Riverside Flight - RC-8-1-89-2005 -- (RCFCWCD) 89 images
- 2005 County of Riverside Flight - RC-9-1-90-2005 -- (RCFCWCD) 88 images
- 2005 County of Riverside Flight - RC-10-1-98-2005 -- (RCFCWCD) 98 images
- 2005 County of Riverside Flight - RC-11-1-88-2005 -- (RCFCWCD) 89 images
- 2005 County of Riverside Flight - RC-12-1-95-2005 -- (RCFCWCD) 95 images
- 2005 County of Riverside Flight - RC-13-1-93-2005 -- (RCFCWCD) 93 images
- 2005 County of Riverside Flight - RC-14-1-98-2005 -- (RCFCWCD) 98 images
- 2005 County of Riverside Flight - RC-15-1-96-2005 -- (RCFCWCD) 96 images
- 2005 County of Riverside Flight - RC-16-1-99-2005 -- (RCFCWCD) 99 images
- 2005 County of Riverside Flight - RC-17-1-105-2005 -- (RCFCWCD) 105 images
- 2005 County of Riverside Flight - RC-18-1-99-2005 -- (RCFCWCD) 99 images
- 2005 County of Riverside Flight - RC-19-1-82-2005 -- (RCFCWCD) 82 images
- 2005 County of Riverside Flight - RC-20-1-82-2005 -- (RCFCWCD) 82 images
- 2005 County of Riverside Flight - RC-21-1-78-2005 -- (RCFCWCD) 78 images
- 2010 County of Riverside Flight - RC-1-1-77-2010 -- (RCFCWCD) 77 images
- 2010 County of Riverside Flight - RC-2-1-79-2010 -- (RCFCWCD) 79 images
- 2010 County of Riverside Flight - RC-3-1-82-2010 -- (RCFCWCD) 82 images
- 2010 County of Riverside Flight - RC-4-1-84-2010 -- (RCFCWCD) 83 images
- 2010 County of Riverside Flight - RC-5-1-86-2010 -- (RCFCWCD) 86 images
- 2010 County of Riverside Flight - RC-6-1-93-2010 -- (RCFCWCD) 92 images
- 2010 County of Riverside Flight - RC-7-1-89-2010 -- (RCFCWCD) 90 images
- 2010 County of Riverside Flight - RC-8-1-90-2010 -- (RCFCWCD) 88 images
- 2010 County of Riverside Flight - RC-9-1-96-2010 -- (RCFCWCD) 96 images
- 2010 County of Riverside Flight - RC-10-1-96-2010 -- (RCFCWCD) 96 images
- 2010 County of Riverside Flight - RC-11-1-94-2010 -- (RCFCWCD) 94 images
- 2010 County of Riverside Flight - RC-12-1-93-2010 -- (RCFCWCD) 93 images
- 2010 County of Riverside Flight - RC-13-1-91-2010 -- (RCFCWCD) 91 images
- 2010 County of Riverside Flight - RC-14-1-95-2010 -- (RCFCWCD) 95 images
- 2010 County of Riverside Flight - RC-15-1-100-2010 -- (RCFCWCD) 97 images
- 2010 County of Riverside Flight - RC-16-1-100-2010 -- (RCFCWCD) 100 images
- 2010 County of Riverside Flight - RC-17-1-101-2010 -- (RCFCWCD) 101 images
- 2010 County of Riverside Flight - RC-18-1-104-2010 -- (RCFCWCD) 105 images
- 2010 County of Riverside Flight - RC-19-1-85-2010 -- (RCFCWCD) 85 images
- 2010 County of Riverside Flight - RC-20-1-81-2010 -- (RCFCWCD) 81 images
- 2010 County of Riverside Flight - RC-21-1-81-2010 -- (RCFCWCD) 81 images
-
Temecula (5,602 images)
The cities of Murrieta, Wildomar, and Temecula are inlocated in the southwestern section of Riverside County..
- 1949 Murrieta Book 12A - RC-AXM-F-6F-5-144-1949 -- (RCFCWCD) 21 images
- 1960 Murrieta - RC-26-33 -- (RCFCWCD) 4 images
- 1961 Murrieta Creek - RC-4-12 -- (RCFCWCD) 9 images
- 1962 Temecula - RC-1-7-1962 -- (RCFCWCD) 7 images
- 1962 Temecula - RC-65-81-1962 -- (RCFCWCD) 17 images
- 1964 Murrieta - RC-27-53-1964 -- (RCFCWCD) 27 images
- 1964 Wildomar-Murrieta Oblique - RC-42-81-1964 -- (RCFCWCD) 39 images
- 1967 Temecula - RC-1-62-1967 -- (RCFCWCD) 60 images
- 1967 Temescal Trunk - RC-1-13-1967 -- (RCFCWCD) 13 images
- 1968 Murrieta Hot Springs - RC-9-26-1968 -- (RCFCWCD) 8 images
- 1968 Portola Rd - RC-1-1968 -- (RCFCWCD) 1 images
- 1968 Rancho California - RC-1-30-1968 -- (RCFCWCD) 30 images
- 1968 Rancho California - RC-1-143-1968 -- (RCFCWCD) 119 images
- 1968 Tenaja Rd - RC-5-17-1968 -- (RCFCWCD) 13 images
- 1970 Menifee - RC-2-4-1970 -- (RCFCWCD) 3 images
- 1972 Auld Valley - RC-1-43-1972 -- (RCFCWCD) 43 images
- 1972 Murrieta Creek - RC-Murrieta-1-21-1972 -- (RCFCWCD) 21 images
- 1972 Murrieta Hot Springs - RC-1-8-1972 -- (RCFCWCD) 8 images
- 1973 Auld Valley Reservoir -RC-1-1-4-6-1973 -- (RCFCWCD) 23 images
- 1973 Murrieta Creek - RC-1-26-1973 -- (RCFCWCD) 24 images
- 1973 Murrieta Hot Spring Road -RC-1-19-1973 -- (RCFCWCD) 14 images
- 1978 Bundy Canyon - RC-189-199-1978 -- (RCFCWCD) 11 images
- 1978 Murrieta-Temecula - RC-1-158-1978 -- (RCFCWCD) 11 images
- 1978 Temecula - RC-201-1978 -- (RCFCWCD) 1 image
- 1978 Washington Street Bridge - RC-200-1978 -- (RCFCWCD) 1 image
- 1978 Wildomar - RC-151-188-1978 -- (RCFCWCD) 30 images
- 1979 Wildomar-Murrieta - RC-1-50-1979 -- (RCFCWCD) 50 images
- 1980 Cahuila Creek - RC-1-6-1980 -- (RCFCWCD) 6 images
- 1980 Long Canyon-Temecula -RC-Long-1-15-1980 -- (RCFCWCD) 15 images
- 1980 Murrieta Creek Restoration - RC-1-44-1980 -- (RCFCWCD) 44 images
- 1980 Murrieta Flood Damage - RC-1-57-1980 -- (RCFCWCD) 58 images
- 1980 Murrieta Flood Damage - RC-1-148-1980 -- (RCFCWCD) 147 images
- 1980 Santa Gertrudis Creek - RC-1-15-1980 -- (RCFCWCD) 15 images
- 1980 Temecula - RC-1-93-1980 -- (RCFCWCD) 85 images
- 1980 Temecula Creek - RC-Temecula-1-57-1980 -- (RCFCWCD) 57 images
- 1980 Tule Creek - RC-1-7-1980 -- (RCFCWCD) 7 images
- 1980 Vai Lake -RC-1-3-1980 -- (RCFCWCD) 3 images
- 1980 Warm Springs Creek - RC-1-12-1980 -- (RCFCWCD) 12 images
- 1980 Wildomar Channel - RC-15-27-1980 -- (RCFCWCD) 13 images
- 1980 Wilson Creek - RC-Wilson-1-34-1980 -- (RCFCWCD) 34 images
- 1981 Wildomar Channel Extension - RC-7-12-1981 -- (RCFCWCD) 6 images
- 1982 Rancho California - RC-1-63-1982 -- (RCFCWCD) 62 images
- 1982 Rancho California Ortho - RC-1-27-1982 -- (RCFCWCD) 27 images
- 1982 Wildomar -RC-4-11-1982 -- (RCFCWCD) 7 images
- 1983 Temecula Wolf Valley -RC-34-39-1983 -- (RCFCWCD) 6 images
- 1983 Wildomar -RC-17-33-1983 -- (RCFCWCD) 17 images
- 1984 Wildomar Channel - RC-1-11-1984 -- (RCFCWCD) 11 images
- 1984 Wilson Valley Road - RC-Wilson-1-6-1984 -- (RCFCWCD) 6 images
- 1985 Murrieta Creek - RC-1-50-1985 -- (RCFCWCD) 50 images
- 1985 Ocotillo Storm Drain - RC-Ocotillo-1-10-1985 -- (RCFCWCD) 4 images
- 1986 Anza Creek Channel - RC-1-9-1986 -- (RCFCWCD) 9 images
- 1986 Benton Road -RC-1-48-60-1986 -- (RCFCWCD) 13 images
- 1986 Murrieta Creek - RC-1-19-1986 -- (RCFCWCD) 19 images
- 1987 Murrieta Creek Channel - RC-1-13-1987 -- (RCFCWCD) 13 images
- 1986 Murrieta Road - RC-1-8-1986 -- (RCFCWCD) 8 images
- 1987 Anza - RC-Anza-1-4-1987 -- (RCFCWCD) 4 images
- 1987 Murrieta Creek - RC-1-48-1987 -- (RCFCWCD) 48 images
- 1987 Scott & Washington - RC-Scott-1-24-1987 -- (RCFCWCD) 24 images
- 1987 Tucalota Ortho - RC-1-10-1987 -- (RCFCWCD) 10 images
- 1988 Bundy Canyon Road - RC-1-21-1988 -- (RCFCWCD) 21 images
- 1988 Golden Triangle Moaiac - RC-1-12-1988 -- (RCFCWCD) 12 images
- 1988 Tucalota - RC-1-45-1988 -- (RCFCWCD) 45 images
- 1989 Murrieta Creek - RC-14-29-1989 -- (RCFCWCD) 29 images
- 1989 Murrieta Creek - RC-Murrieta-1-21-1989 -- (RCFCWCD) 20 images
- 1989 Murrieta Creek II - RC-1-36-1989 -- (RCFCWCD) 36 images
- 1989 Murrieta-F - RC-1-7-1989 -- (RCFCWCD) 7 images
- 1989 Murrieta III - RC-1-38-1989 -- (RCFCWCD) 38 images
- 1990 Murrieta Creek - RC-1-13-1990 -- (RCFCWCD) 13 images
- 1991 Murrieta Creek - RC-1-57-1991 -- (RCFCWCD) 58 images
- 1991 Murrieta Line F-1 - RC-Murrieta-1-6-1991 -- (RCFCWCD) 6 images
- 1991 Riverside / San Bernardino 1991 - RC-M-22-S-29-M -- (RCFCWCD) 11 images
- 1991 Riverside / San Bernardino 1991 - RC-M-22-S-29-N -- (RCFCWCD) 10 images
- 1991 Riverside / San Bernardino 1991 - RC-M-22-S-29-O -- (RCFCWCD) 6 images
- 1991 Riverside / San Bernardino 1991 - RC-M-22-S-29-P -- (RCFCWCD) 6 images
- 1991 Riverside / San Bernardino 1991 - RC-M-22-S-29-Q -- (RCFCWCD) 4 images
- 1991 Riverside / San Bernardino 1991 - RC-M-22-S-29-R -- (RCFCWCD) 3 images
- 1991 Riverside / San Bernardino 1991 - RC-M-22-S-29-S -- (RCFCWCD) 3 images
- 1991 Santa Gertrudis Creek - RC-1-11-1991 -- (RCFCWCD) 11 images
- 1991 Wildomar Channel - RC-1-13-1991 -- (RCFCWCD) 13 images
- 1991 Wildomar Lateral C -RC-Wildomar-1A-3B-1991 -- (RCFCWCD) 6 images
- 1991 Wildomar Lateral E - RC-Wildomar-1A-2B-1991 -- (RCFCWCD) 4 images
- 1993 Murrieta - RC-1-84-1993 -- (RCFCWCD) 84 images
- 1993 Murrieta Creek Restoration - RC-1-39-1993 -- (RCFCWCD) 39 images
- 1994 Murrieta Basin - RC-1-19-1994 -- (RCFCWCD) 19 images
- 1994 Murrieta Creek Restoration - RC-1-39-1994 -- (RCFCWCD) 39 images
- 1994 Murrieta Line A - RC-Murrieta-1-2-1994 -- (RCFCWCD) 2 images
- 1993 Murrieta-Temecula - RC-1-307-1993-1 -- (RCFCWCD) 251 images
- 1993 Murrieta-Temecula - RC-1-307-1993-2 -- (RCFCWCD) 58 images
- 1994 Murrieta Creek - RC-1-118-1994 -- (RCFCWCD) 118 images
- 1994 Murietta Ortho - RC-1-57-1994 -- (RCFCWCD) 59 images
- 1994 Santa Gertrudis Creek - RC-1-40-1994 -- (RCFCWCD) 40 images
- 1994 Tucalota Creek - RC-Tucalota-1-6-1994 -- (RCFCWCD) 5 images
- 1994 Tucalota Creek - RC-1-1994 -- (RCFCWCD) 1 image
- 1994 Warm Springs Channel - RC-Warm-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Wildomar Channel - RC-Wildomar-1-4-1994 -- (RCFCWCD) 4 images
- 1994 Wildomar "C" - RC-Wildomar-1-5-1994 -- (RCFCWCD) 58 images
- 1995 Anza Sanitary Landfill - RC-Anza2-1-3-1995 -- (RCFCWCD) 3 images
- 1995 Anza Sanitary Landfill Spot - RC-Anza-A-1995 -- (RCFCWCD) 1 image
- 1995 Murrieta - RC-1-54-1995 -- (RCFCWCD) 54 images
- 1995 Murrieta Creek Channel - RC-1-88-1995 -- (RCFCWCD) 88 images
- 1995 Murrieta Ortho Phase II - RC-1A-4A-1995 -- (RCFCWCD) 4 images
- 1995 Temecula Creek - RC-1-15-1995 -- (RCFCWCD) 15 images
- 1996 FEMA Revised Murrieta Line F - RC-Fema-1-8-1996 -- (RCFCWCD) 8 images
- 1996 Lake Skinner Ortho - RC-1-51-1996 -- (RCFCWCD) 51 images
- 1996 Murrieta Ortho Phase III - RC-Murrieta-1-41-1996 -- (RCFCWCD) 41 images
- 1996 Temecula Creek - RC-Temecula-1-16-1996 -- (RCFCWCD) 16 images
- 1996 Wildomar Channel - RC-Wildomar-1-8-1996 -- (RCFCWCD) 8 images
- 1997 Murrieta Creek - RC-1-68-1997 -- (RCFCWCD) 68 images
- 1997 Murrieta Creek - RC-Murrieta2-1-72-1997 -- (RCFCWCD) 69 images
- 1997 Murrieta Creek Line D and D1 - RC-10-20-1997 -- (RCFCWCD) 11 images
- 1997 Murrieta Creek Line D and D1 - RC-Murrieta-1-9-1997 -- (RCFCWCD) 9 images
- 1997 Murrieta Creek Lines F and F1 - RC-1-19-1997 -- (RCFCWCD) 19 images
- 1997 Murrieta Creek Stage 1 Update Area 1 - RC-1-16-1997 -- (RCFCWCD) 26 images
- 1997 Murrieta Creek Stage 1 Update Area 2 - RC-1-6-1997 -- (RCFCWCD) 6 images
- 1997 Murrieta Line F and F1 - RC-Murrieta-1-16-1997 -- (RCFCWCD) 16 images
- 1997 Sage Road - RC-1-21-1997 -- (RCFCWCD) 21 images
- 1997 Santa Gertrudis Creek - RC-Gertrudis-1-13-1997 -- (RCFCWCD) 13 images
- 1997 Wildomar Channel - RC-Wildomar-1-16-1997 -- (RCFCWCD) 13 images
- 1997 Wildomar Channel Lateral C - RC-Wildomar2-1-6-1997 -- (RCFCWCD) 6 images
- 1999 Murrieta Creek - RC-1-68-1999 -- (RCFCWCD) 68 images
- 1999 Murrieta Creek - RC-1-73-2000 -- (RCFCWCD) 73 images
- 1999 Murrieta F & F1 - RC-Murrieta-1-17-1999 -- (RCFCWCD) 17 images
- 1999 Murrieta Main Drain Pipe Line - RC-Murrieta-1-9-1999 -- (RCFCWCD) 9 images
- 1999 Murrieta Main Drain Pipe Line F-1 and F - RC-Murrieta-1-16-1999 -- (RCFCWCD) 16 images
- 1999 Santa Gertrudis Creek - RC-1-20-1999 -- (RCFCWCD) 20 images
- 1999 Wildomar Channel - RC-Wildomar2-1-18-1999 -- (RCFCWCD) 18 images
- 1999 Wildomar Channel - RC-1-6-1999 -- (RCFCWCD) 6 images
- 1999 Wildomar Channel - RC-1-19-1999 -- (RCFCWCD) 19 images
- 1999 Wildomar Channel Lateral C - RC-Wildomar-1-7-1999 -- (RCFCWCD) 7 image
- 2000 Murrieta Creek - RC-1-73-2000 -- (RCFCWCD) 73 images
- 2000 Murrieta Creek Line D - RC-Murrieta-1-3-2000 -- (RCFCWCD) 3 images
- 2000 Murrieta F & F1 - RC-Murrieta-1-17-2000 -- (RCFCWCD) 17 images
- 2000 Santa Gertrudis Creek - RC-1-23-2000 -- (RCFCWCD) 23 images
- 2000 Wildomar Channel - RC-Wildomar-1-17-2000 -- (RCFCWCD) 17 images
- 2000 Wildomar Channel Lateral C - RC-WildomarC-1-6-2000 -- (RCFCWCD) 6 images
- 2001 Murrieta Line F and F1 - RC-1-16-2001 -- (RCFCWCD) 16 images
- 2001 Murrieta Creek - RC-1-69-2001 -- (RCFCWCD) 69 images
- 2001 Santa Gertrudis Creek - RC-1-22-2001 -- (RCFCWCD) 22 images
- 2001 Wildomar Channel - RC-Wildomar-1-17-2001 -- (RCFCWCD) 17 images
- 2001 Wildomar Lateral C - RC-Wildomar2-1-6-2001 -- (RCFCWCD) 6 images
- 2002 Murrieta Interim Basin - RC-Murrieta-1-9-2002 -- (RCFCWCD) 9 images
- 2003 Murrieta Creek - RC-1-58-2003 -- (RCFCWCD) 58 images
- 2003 Murrieta Creek - RC-1-48-2003 -- (RCFCWCD) 48 images
- 2003 Murrieta Creek - RC-Murrieta3-1-8-2003 -- (RCFCWCD) 8 images
- 2003 Murrieta Creek Line F - RC-Murrieta-1-9-2003 -- (RCFCWCD) 9 images
- 2003 Murrieta Creek Line F - RC-1-9-2003 -- (RCFCWCD) 9 images
- 2003 Murrieta Creek Obliques - RC-Murrieta2-1-3-2003 -- (RCFCWCD) 3 images
- 2003 Murrieta Creek Spot - RC-Murrieta-A-2003 -- (RCFCWCD) 1 image
- 2003 Murrieta Valley I-215 Basin - RC-Murrieta-1-2-2003 -- (RCFCWCD) 2 images
- 2003 Santa Gertrudis - RC-1-21-2003 -- (RCFCWCD) 21 images
- 2003 Tucolata Creek - RC-1-8-2003 -- (RCFCWCD) 8 images
- 2003 Wildomar Channel Main Drain Pipe - RC-1-14-2003 -- (RCFCWCD) 14 images
- 2003 Wildomar Main Drain Pipe Lateral C - RC-Wildomar-1-5-2003 -- (RCFCWCD) 5 images
- 2004 Murrieta Creek - RC-1-50-2004 -- (RCFCWCD) 50 images
- 2004 Murrieta Creek - RC-A-2004 -- (RCFCWCD) 1 image
- 2004 Murrieta Creek - RC-B-2004 -- (RCFCWCD) 1 image
- 2004 Murrieta Interim Basin - RC-Murrieta-1-9-2004 -- (RCFCWCD) 9 images
- 2004 Murrieta Creek - RC-Murrieta2-1-7-2004 -- (RCFCWCD) 7 images
- 2004 Santa Gertrudis Creek - RC-1-22-2004 -- (RCFCWCD) 22 images
- 2004 Tucolata Creek - RC-1-8-2004 -- (RCFCWCD) 8 images
- 2004 Wildomar Channel - RC-1-13-2004 -- (RCFCWCD) 13 images
- 2005 Menifee - RC-1-89-2005 -- (RCFCWCD) 89 images
- 2005 Murrieta Creek - RC-Murrieta7-1-48-2005 -- (RCFCWCD) 47 images
- 2005 Murrieta Creek - RC-1-1-2-12-2005 -- (RCFCWCD) 22 images
- 2005 Murrieta Creek Line F - RC-1-9-2005 -- (RCFCWCD) 9 images
- 2005 Murrieta Creek - RC-1-49-2005 -- (RCFCWCD) 49 images
- 2005 Murrieta Interim Basin - RC-Murrieta2-1-9-2005 -- (RCFCWCD) 9 images
- 2005 Murrieta Valley I-215 Basin - RC-Murrieta-1-2-2005 -- (RCFCWCD) 2 images
- 2005 Santa Gertrudic Creek - RC-1-21-2005 -- (RCFCWCD) 21 images
- 2005 Southwest Menifee - RC-1-8-2005 -- (RCFCWCD) 8 images
- 2005 Temecula Creek - RC-1-1-2-6-2005 -- (RCFCWCD) 6 images
- 2005 Temecula Creek - RC-1-12-2005 -- (RCFCWCD) 6 images
- 2005 Temecula Ortho - RC-1-24-2005 -- (RCFCWCD) 24 images
- 2005 Tucolata Creek - RC-Tucolata-1-8-2005 -- (RCFCWCD) 8 images
- 2005 Wildomar Channel - RC-Wildomar-1-13-2005 -- (RCFCWCD) 13 images
- 2006 Murrieta Creek -RC-Murrieta6-1-2006 -- (RCFCWCD) 1 image
- 2006 Murrieta Creek - RC-Murrieta7-1-5-2006 -- (RCFCWCD) 5 images
- 2006 Murrieta Creek Line F - RC-Murrieta3-1-9-2006 -- (RCFCWCD) 9 images
- 2006 Murrieta Interim Basin - RC-Murrieta9-1-8-2006 -- (RCFCWCD) 8 images
- 2006 Murrieta Interim Basin - RC-Murrieta2-1-9-2006 -- (RCFCWCD) 9 images
- 2006 Murrieta Valley I-215 Basin - RC-Murrieta8-1-2-2006 -- (RCFCWCD) 8 images
- 2006 Murrieta Valley I-215 Basin - RC-Murrieta-1-2-2006 -- (RCFCWCD) 2 images
- 2006 Santa Gertrudis Creek - RC-1-21-2006 -- (RCFCWCD) 21 images
- 2006 Santa Gertrudis Creek - RC-1-21-2006 -- (RCFCWCD) 21 images
- 2006 Tucolate Creek - RC-Tucolate6-1-8-2006 -- (RCFCWCD) 8 images
- 2006 Tucolata Creek - RC-Tucolata-1-8-2006 -- (RCFCWCD) 8 images
- 2006 Wildomar Channel - RC-1-13-2006 -- (RCFCWCD) 13 images
- 2007 Murrieta Creek Line F - RC-1-9-2007 -- (RCFCWCD) 9 images
- 2007 Murrieta Creek - RC-1-20-2007 -- (RCFCWCD) 20 images
- 2007 Murrieta Creek - RC-1-48-2007 -- (RCFCWCD) 49 images
- 2007 Wildomar Channel - RC-1-13-2007 -- (RCFCWCD) 13 images
- 2007 Wildomar Channel Lateral B Debris Basin - RC-1-2-2007 -- (RCFCWCD) 2 images
- 2008 Murrieta Creek - RC-Murrieta5-1-48-2008 -- (RCFCWCD) 48 images
- 2008 Murrieta Creek - RC-1-48-2008 -- (RCFCWCD) 48 images
- 2008 Murrieta Creek Channel Interim Basin - RC-Murrieta9-1-9-2008 -- (RCFCWCD) 9 images
- 2008 Murrieta Creek Line F - RC-Murrieta6-1-9-2008 -- (RCFCWCD) 9 images
- 2008 Murrieta Creek Line F - RC-Murrieta8-1-9-2008 -- (RCFCWCD) 9 images
- 2008 Murrieta Interim Basin - RC-Murrieta4-1-9-2008 -- (RCFCWCD) 9 images
- 2008 Murrieta Valley I-215 Basin - RC-Murrieta5-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Murrieta Valley I-215 Basin - RC-Murrieta7-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Pechanga Creek - RC-Pechanga7-1-7-2008 -- (RCFCWCD) 7 images
- 2008 Pechanga Creek - RC-Pechanga-1-7-2008 -- (RCFCWCD) 7 images
- 2008 Santa Gertrudis Creek - RC-1-21-2008 -- (RCFCWCD) 21 images
- 2008 Santa Gertrudis Creek - RC-Santa-1-21-2008 -- (RCFCWCD) 21 images
- 2008 Temecula Creek - RC-Temecula7-1-35-2008 -- (RCFCWCD) 37 images
- 2008 Temecula Channel - RC-1-37-2008 -- (RCFCWCD) 37 images
- 2008 Tucolata Creek - RC-Tucolata8-1-8-2008 -- (RCFCWCD) 8 images
- 2008 Tucolata - RC-Tucolata-1-8-2008 -- (RCFCWCD) 8 images
- 2008 Wildomar Channel - RC-Wildomar5-1-13-2008 -- (RCFCWCD) 13 images
- 2008 Wildomar Channel - RC-Wildomar7-1-13-2008 -- (RCFCWCD 13 images
- 2008 Wildomar Channel Lateral B - RC-Wildomar6-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Wildomar Channel Lateral B Debris Basin - RC-Wildomar8-1-2-2008 -- (RCFCWCD) 2 images
- 2008 Wildomar Channel Lateral C - RC-Wildomar4-1-7-2008 -- (RCFCWCD) 7 images
- 2008 Wildomar Lateral C, Stage 3 - RC-Wildomar9-1-7-2008 -- (RCFCWCD) 7 images
- 2009 Murrieta Valley I-215 - RC-Murrieta-1-2-2009 -- (RCFCWCD) 2 images
- 2009 Pechanga Creek - RC-Pechanga-1-8-2009 -- (RCFCWCD) 8 images
- 2009 Wildomar Channel - RC-Wildomar-1-4-2009 -- (RCFCWCD) 4 images
- 2010 Murrieta Creek - RC-1-26-2010 -- (RCFCWCD) 26 images
- 2010 Murrieta Creek Line F - RC-1-9-2010 -- (RCFCWCD) 9 images
- 2010 Santa Gertrudis Creek - RC-1-22-2010 -- (RCFCWCD) 22 images
- 2010 Tucolata Creek - RC-Tucolata-1-8-2010 -- (RCFCWCD) 8 images
- 2010 Wildomar Channel Lateral A - RC-1-2-2010 -- (RCFCWCD) 2 images
- 2010 Wildomar Channel Lateral C - RC-1-7-2010 -- (RCFCWCD) 7 images
- 2011 Murrieta Creek - RC-1-48-2011 -- (RCFCWCD) 48 images
- 2011 Murrieta Interim Basin - RC-Murrieta2-1-9-2011 -- (RCFCWCD) 9 images
- 2011 Murrieta Valley Creek Line F - RC-MurrietaF-1-9-2011 -- (RCFCWCD) 9 images
- 2011 Murrieta Valley I-215 Basin - RC-Murrieta-1-2-2011-- (RCFCWCD) 2 images
- 2011 Pechanga Creek - RC-Pechanga-1-7-2011 -- (RCFCWCD) 7 images
- 2011 Santa Gertrudis Creek - RC-1-21-2011 -- (RCFCWCD) 21 images
- 2011 Temecula Channel - RC-1-37-2011 -- (RCFCWCD) 37 images
- 2011 Tucolata Creek - RC-Tucolata-1-8-2011 -- (RCFCWCD) 8 images
- 2011 Wildomar Canyon - RC-1-13-2011 -- (RCFCWCD) 13 images
- 2011 Wildomar Lateral B - RC-WildomarB-1-2-2011 -- (RCFCWCD) 2 images
- 2011 Wildomar Lateral C, Stage 3 - RC-WildomarC-1-7-2011 -- (RCFCWCD) 7 images
- 2012 Agriscape Lakeview Murrieta - RC-AG-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Murrieta Creek - RC-1-48-2012 -- (RCFCWCD) 48 images
- 2012 Murrieta Creek Line F - RC-MurrietaF-1-9-2012 -- (RCFCWCD) 9 images
- 2012 Murrieta Interim Basin - RC-Murrieta3-1-9-2012 -- (RCFCWCD) 9 images
- 2012 Murrieta Valley I-215 - RC-Murrieta2-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Pechanga Creek - RC-Pechanga-1-7-2012 -- (RCFCWCD) 7 images
- 2012 Santa Gertrudis Creek - RC-1-21-2012 -- (RCFCWCD) 21 images
- 2012 Temecula Channel - RC-1-37-2012 -- (RCFCWCD) 37 images
- 2012 Tucolata Creek - RC-Tucolata-1-8-2012 -- (RCFCWCD) 8 images
- 2012 Wildoma Channel -RC-1-13-2012 -- (RCFCWCD) 13 images
- 2012 Wildomar Channel Lateral B Debris Basin- RC-Wildomar-1-2-2012 -- (RCFCWCD) 2 images
- 2012 Wildomar Lateral C, Stage 3-RC-WildomarC-1-7-2012 -- (RCFCWCD) 7 images
- 2013 Murrieta Creek - RC-1-48-2013 -- (RCFCWCD) 48 images
- 2013 Murrieta Interior Basin - RC-Murrieta2-1-9-2013 -- (RCFCWCD) 9 images
- 2013 Murrieta Line F - RC-MurrietaF-1-9-2013 -- (RCFCWCD) 9 images
- 2013 Murrieta Valley I-215 Basin - RC-Murrieta-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Pechanga Creek - RC-Pechanga-1-7-2013 -- (RCFCWCD) 7 images
- 2013 Santa Gertrudis Creek - RC-1-21-2013 -- (RCFCWCD) 21 images
- 2013 Temecula Channel - RC-1-37-2013 -- (RCFCWCD) 37 images
- 2013 Tucolata Creek - RC-Tucolata-1-8-2013 -- (RCFCWCD) 8 images
- 2013 Wildomar Channel - RC-Wildomar2-1-13-2013 -- (RCFCWCD) 13 images
- 2013 Wildomar Channel Lateral B Debris Basin - RC-WildomarB-1-2-2013 -- (RCFCWCD) 2 images
- 2013 Wildomar Lateral C Stage 3 -RC-Wildomar3-1-7-2013 -- (RCFCWCD) 7 images
- 2014 Murrieta Valley I-215 Basin - RC-Murrieta-1-2-2014 -- (RCFCWCD) 2 images
- 2014 Pechanga Creek - RC-1-7-2014 -- (RCFCWCD) 7 images
- 2014 Santa Gertrudis Creek - RC-1-21-2014 -- (RCFCWCD) 21 images
- 2014 Temecula Channel - RC-1-37-2014 -- (RCFCWCD) 37 images
- 2014 Tucolata Creek - RC-1-8-2014 -- (RCFCWCD) 8 images
- 2015 Murrieta Creek - RC-1-48-2015 -- (RCFCWCD) 48 images
- 2015 Murrieta Creek Line F - RC-MurrietaF-1-9-2015 -- (RCFCWCD) 9 images
- 2015 Murrieta Interim Basin - RC-Murrieta-1-9-2015 -- (RCFCWCD) 9 images
- 2015 Murrieta Valley I-215 Basin - RC-Murrieta215-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Pechanga Creek - RC-1-7-2015 -- (RCFCWCD) 7 images
- 2015 Santa Gertrudis Creek - RC-1-21-2015 -- (RCFCWCD) 21 images
- 2015 Temecula Channel - RC-1-37-2015 -- (RCFCWCD) 37 images
- 2015 Tucolata Creek - RC-Tucolata-1-8-2015 -- (RCFCWCD) 8 images
- 2015 Wildomar Channel- RC-Wildomar-1-13-2015 -- (RCFCWCD) 14 images
- 2015 Wildomar Channel Lateral B Debris Basin - RC-WildomarB-1-2-2015 -- (RCFCWCD) 2 images
- 2015 Wildomr Lateral C Stage 3 - RC-WildomarC-1-7-2015 -- (RCFCWCD) 7 images
Sacramento Delta (42 Images)
- 1949 Delta -- Various Flights (Rowe) 42 images
San Bernardino County (32,677 Images)
-
USDA Photographs of San Bernardino County (10,874 images)
The USDA photographed San Bernardino County in the early 1950's. The pictures are organized according to photographic sheets that represent a sector of the County. Each Index Sheet is created by using individual photos in an overlapping mosaic to show a larger geographic area. Each individul photo that made up this mosaic may look quite different alone as only a small portion of the photo may have been used to create the entire section.
The location of each particular geographic section is a general description. The entire flight covers a very wide area and some locations are quite remote. San Bernardino County sections that are available for viewing range from the Western San Bernardino County to the Eastern San Bernardino County. When you locate the section of interest, the number corresponds to the USDA Index Sheets below.
Note: Each reference below includes a picture of the USDA-SBCFCD index sheet at the start of the reference and individual pictures at the end of the reference (pictures). All USDA pictures in this sector of San Bernardino County are included on the index sheet. Each picture has an identifying number if you want to locate a specific photo. To enlarge the index sheet photograph for viewing in greater detail, you should move your cursor over the area of interest and click the mouse. If you want smaller increase in size, you should press Ctrl + "+" if on a PC computer or press CMD + "+" if on a MacIntosh computer. To reduce the magnification of the photograph, click the mouse over the picture. If you want a smaller decrease in size, press Ctrl + "-" if on a PC computer or press CMD + "-" if on a MacIntosh computer.
- USDA Index Sheet 1 (Mojave Desert near South Inyo County in the area of Death Valley Road and State Route 127) - pictures 117 images
- USDA Index Sheet 2 (Mojave Desert near Fort Irwin) - pictures 209 images
- USDA Index Sheet 3 (Mojave Desert south of Death Valley National Park) - pictures 206 images
- USDA Index Sheet 4 (Mojave Desert south of Death Valley National Park) - pictures 104 images
- USDA Index Sheet 5 (Mojave Desert near Trona) - pictures 178 images
- USDA Index Sheet 6 (Mojave Desert near China Lake) - pictures 143 images
- USDA Index Sheet 7 (Mojave Desert near Red Mountain) - pictures 73 images
- USDA Index Sheet 8 (Mojave Desert near Fort Irwin) - pictures 157 images
- USDA Index Sheet 9 (Mojave Desert near Fort Irwin) - pictures 101 images
- USDA Index Sheet 10 (Mojave Desert near Fort Irwin & Bicycle Lake) - pictures 181 images
- USDA Index Sheet 11 (Mojave Desert west of Baker) - pictures 104 images
- USDA Index Sheet 12 (Mojave Desert near Baker) - pictures 122 images
- USDA Index Sheet 13 (Mojave Desert south of Baker and Interstate 15) - pictures 149 images
- USDA Index Sheet 14 (Mojave Desert southwest of Baker and Interstate 15) - pictures 189 images
- USDA Index Sheet 15 (Mojave River Watershed near Yermo) - pictures 124 images
- USDA Index Sheet 16 (Mojave Desert north of Barstow) - pictures 75 images
- USDA Index Sheet 17 (Mojave River Watershed near Yermo, Newberry Springs, & Barstow) - pictures 128 images
- USDA Index Sheet 18 (Mojave Desert near Kramer Junction & Boron) - pictures 64 images
- USDA Index Sheet 19 (Mojave River Watershed near Adelanto) - pictures 74 images
- USDA Index Sheet 20 (Mojave River Watershed near Silverlakes &Helendale) - pictures 138 images
- USDA Index Sheet 21 (Mojave River Watershed near Hinkley, Newberry Springs, & Barstow) - pictures 82 images
- USDA Index Sheet 22 (Mojave River Watershed near Yermo, Newberry Springs, & Dagget) - pictures 196 images
- USDA Index Sheet 23 (Mojave River Watershed near Barstow) - pictures 192 images
- USDA Index Sheet 24 (Mojave Desert near Ludlow) - pictures 174 images
- USDA Index Sheet 25 (Mojave Desert near Bagdad) - pictures 73 images
- USDA Index Sheet 26 (Mojave Desert near 29 Palms) - pictures 59 images
- USDA Index Sheet 27 (Mojave Desert near 29 Palms) - pictures 119 images
- USDA Index Sheet 28 (Mojave Desert near Landers) - pictures 182 images
- USDA Index Sheet 29 (Mojave Desert north of 29 Palms) - pictures 170 images
- USDA Index Sheet 30 (Mojave River Watershed near Apple Valley & Lucerne Valley) - pictures 117 images
- USDA Index Sheet 31 (Mojave River Watershed near Apple Valley & Victorville) - pictures 120 images
- USDA Index Sheet 32 (Mojave Desert near Phelan) - pictures 116 images
- USDA Index Sheet 33 (Santa Ana Watershed near Lytle Creek, Wrightwood, Upland & San Antonio Heights) - pictures 118 images
- USDA Index Sheet 34 (Mojave River Watershed & Santa Ana Watershed near Lytle Creek, Hesperia & Cajon Pass, ) - pictures 148 images
- USDA Index Sheet 35 (Mojave River Watershed & Santa Ana Watershed near Lake Arrowhead, Big Bear Lake, Lucerne USDA Index Valley & Apple Valley) - pictures 196 images
- USDA Index Sheet 36 (Santa Ana Watershed near Chino, Ontario, Corona, Prado Dam, Santa Ana River, Chino Hills, Pomona & Fontana) - pictures 98 images
- USDA Index Sheet 37 (Santa Ana Watershed near San Bernardino, Colton, Redlands, Lytle Creek, Bloomington & Rialto) - pictures 90 images
- USDA Index Sheet 38 (Santa Ana Watershed near Redlands, Yucaipa, Mentone, Highland, City Creek & Mill Creek) - pictures 96 images
- USDA Index Sheet 39 (Near San Bernardino National Forest, Barton Flats, Big Bear, Lake Baldwin & Jenks Lake) - pictures 183 images
- USDA Index Sheet 40 (Near Yucca Valley, Joshua Tree & Pioneer Town) - pictures 133 images
- USDA Index Sheet 41 (Near 29 Palms) - pictures 161 images
- USDA Index Sheet 42 (Near 29 Palms) - pictures 103 images
- USDA Index Sheet 43 (Near Amboy & Cadiz) - pictures 86 images
- USDA Index Sheet 44 (Near Amboy & Cadiz) - pictures 87 images
- USDA Index Sheet 45 (Near Essex) - pictures 107 images
- USDA Index Sheet 46 (Near Goffs) - pictures 70 images
- USDA Index Sheet 47 (Near Goffs) - pictures 134 images
- USDA Index Sheet 48 (Near Ward Valley) - pictures 55 images
- USDA Index Sheet 49 (Mojave Desert near the Colorado River north of Needles) - pictures 96 images
- USDA Index Sheet 50 (Lower Colorado River Watershed near Needles & Mohave Valley) - pictures 110 images
- USDA Index Sheet 51 (Lower Colorado River Watershed near Mojave Desert, Needles, Topock, Lower Colorado River & Mohave Valley) - pictures 198 images
- USDA Index Sheet 52 (Near Needles airport) - pictures 161 images
- USDA Index Sheet 53 (Lower Colorado River Watershed near Lower Colorado River, Lake Havasu & Lake Havasu City) - USDA Index pictures 93 images
- USDA Index Sheet 54 (Near Vidal Junction, Vidal & Colorado River Aqueduct) - pictures 101 images
- USDA Index Sheet 55 (Lower Colorado Watershed near Parker, Parker Dam, Lower Colorado River, Earp & Big River) - pictures 119 images
- USDA Index Sheet 56 (Lower Colorado Watershed at the Arizona-California boundary) - pictures 101 images
- USDA Index Sheet 57 (Mojave River Watershed near Newberry Springs ) - pictures 138 images
- USDA Index Sheet 58 (Mojave River Watershed near Barstow ) - pictures 96 images
- USDA Index Sheet 59 (Mojave River Watershed near Barstow and Lynwood ) - pictures 116 images
- USDA Index Sheet 60 (Mojave River Watershed near Victorville, George AFB) - pictures 132 images
- USDA Index Sheet 61 (Near Phelan ) - pictures 146 images
- USDA Index Sheet 62 (Mojave River Watershed near Victorville and Hesperia) - pictures 107 images
- USDA Index Sheet 63 (Near Hesperia) - pictures 114 images
- USDA Index Sheet 64 (Santa Ana Watershed near Redlands, Yucaipa, including San Bernardino National Forest ) - pictures 134 images
- USDA Index Sheet 65 (Santa Ana Watershed near San Bernardino, Colton and Fontana ) - pictures 173 images
- USDA Index Sheet 66 (Santa Ana Watershed near Ontario and Upland) - pictures 135 images
- USDA Index Sheet 67 (Santa Ana Watershed near Chino and Prado Dam ) - pictures 33 images
- USDA Index Sheet 68 (Santa Ana Watershed near San Bernardino, Redlands, including San Bernardino National Forest ) - pictures 131 images
- USDA Index Sheet 69 (Santa Ana Watershed near Redlands, Mentone, including San Bernardino National Forest) - pictures 104 images
- USDA Index Sheet 70 (Mojave River Watershed near Victorville, including San Bernardino National Forest ) - pictures 109 images
- USDA Index Sheet 71 (Mojave River Watershed near Apple Valley ) - pictures 136 images
- USDA Index Sheet 72 (Victorville, Mojave River Watershed near Victorville and Silver Lakes ) - pictures 117 images
- USDA Index Sheet 73 (Mojave River Watershed near Lynwood and Hinkley )- pictures 103 images
- USDA Index Sheet 74 (Mojave River Watershed near Newberry Springs ) - pictures 60 images
- USDA Index Sheet 75 (Mojave River Watershed near Newberry Springs and Yermo ) - pictures 145 images
- USDA Index Sheet 76 (Lower Colorado River Watershed near Fort Mojave ) - pictures 8 images
-
29 Palms (2,704 Images)
Cities that are included in this section include Apple Valley, Deep Creek, Helendale, Hesperia, Lucerne Valley, Oro Grande, Spring Valley Lake, Stoddard Wells, and Victorville.
- 1942 Pine Knot - Flight F-32-P31 -- (SBCFCD) 3 images 1942 Pine Knot - Flight F-32-P31 Index
- 1942 Pine Knot - Flight F-32-P32 -- (SBCFCD) 3 images 1942 Pine Knot - Flight F-32-P32 Index
- 1942 Pine Knot - Flight F-32-P33 -- (SBCFCD) 4 images 1942 Pine Knot - Flight F-32-P33 Index
- 1942 Pine Knot - Flight F-32-P34 -- (SBCFCD) 5 images Pine Knot - Flight F-32-P34 Index
- 1942 Pine Knot - Flight F-32-P35 -- (SBCFCD) 4 images 1942 Pine Knot - Flight F-32-P35 Index
- 1960 Coyote Creek - Flight C31 -- (SBCFCD) 8 images 1960 Coyote Creek - Flight C31 Index
- 1964 29 Palms - Flight E-51 -- (SBCFCD) 86 images 1964 29 Palms - Flight E-51 Index
- 1964 29 Palms - Flight E-52 -- (SBCFCD) 71 images 964 29 Palms - Flight E-52 Index
- 1964 29 Palms - Flight E-54 -- (SBCFCD) 93 images 1964 29 Palms - Flight E-54 Index
- 1964 29 Palms - Flight E-55 -- (SBCFCD) 114 images 1964 29 Palms - Flight E-55 Index
- 1964 Copper Mountain - Flight E-48 -- (SBCFCD) 102 images 1964 Copper Mountain - Flight E-48 Index
- 1964 Dry Coller Lake - Flight E-47 -- (SBCFCD) 92 images 1964 Dry Coller Lake - Flight E-47 Index
- 1964 East Desert - Dale Lake - Flight E-57 -- (SBCFCD) 195 images 1964 East Desert - Dale Lake - Flight E-57 Index
- 1964 East Desert - Hum bug_Mnt - Flight E-58 -- (SBCFCD) 116 images 1964 East Desert - Hum bug_Mnt - Flight E-58 Index
- 1964 East Desert Morongo - Flight E-43 -- (SBCFCD) 43 images 1964 East Desert Morongo - Flight E-43 Index
- 1964 East Desert Morongo - Flight E-44 -- (SBCFCD) 80 images 1964 East Desert Morongo - Flight E-44 Index
- 1964 East Desert Morongo - Flight E-45 -- (SBCFCD) 95 images 1964 East Desert Morongo - Flight E-45 Index
- 1964 East Desert - Sheep Peak - Flight E-56 -- (SBCFCD) 57 images 1964 East Desert - Sheep Peak - Flight E-56 Index
- 1964 East Desert - Valley Mnt - Flight E-53 -- (SBCFCD) 102 images 1964 East Desert - Valley Mnt - Flight E-53 Index
- 1964 East Valley Morongo - Flight E-41 -- (SBCFCD) 1 image 1964 East Valley Morongo - Flight E-41 Index
- 1964 Joshua Tree - Flight E-49 -- (SBCFCD) 37 images 1964 Joshua Tree - Flight E-49 Index
- 1964 Mesquite Lake - Flight E-50 -- (SBCFCD) 95 images 1964 Mesquite Lake - Flight E-50 Index
- 1964 Yucca Valley - Flight E-46 -- (SBCFCD) 102 images 1964 Yucca Valley - Flight E-46 Index
- 1967 Copper Mountain Park - Flight C166 -- (SBCFCD) 3 images Index
- 1967 29 Palms - Pinto Cover - Flight C135 -- (SBCFCD) 20 images Index
- 1967 Refly of #40 - Flight C134 -- (SBCFCD) 28 images Index
- 1968 Yucca Valley - Flight C152 -- (SBCFCD) 19 images Index
- 1968 Yucca Valley - Flight C161 -- (SBCFCD) 18 images Index
- 1971 Yucca Creek - Flight C181 -- (SBCFCD) 20 images Index
- 1972 29 Palms / Utah Trail - Flight C191 -- (SBCFCD) 7 images
- 1972 29 Palms - Flight 220 -- (SBCFCD) 3 images
- 1972 Joshua Tree - Flight C212 -- (SBCFCD) 6 images
- 1972 Morongo Valley - Flight C198 -- (SBCFCD) 11 images Index
- 1973 29 Palms - Flight C222 -- (SBCFCD) 8 images
- 1973 Morongo - Flight C215 -- (SBCFCD) 2 images
- 1974 29 Palms Disposal Site - Flight C249 -- (SBCFCD) 3 images
- 1976 29 Palms - Flight C384 -- (SBCFCD) 1 image
- 1977 29 Palms - Flight C277 -- (SBCFCD) 51 images Index
- 1977 29 Palms - Flight 278 -- (SBCFCD) 52 images Index
- 1977 East Desert - Flight C316 -- (SBCFCD) 4 images Index
- 1977 Quail Wash - Flight C276 -- (SBCFCD) 8 images Index
- 1977 Wonder Valley - Flight C284 -- (SBCFCD) 41 images Index
- 1979 Morongo - Yucca Valley - Flight C291 -- (SBCFCD) 98 images Index
- 1979 Yucca Valley-Old Woman Springs - Flight C416 -- (SBCFCD) 33 images
- 1980 Morongo Valley - Flight C319 -- (SBCFCD) 48 images Index
- 1983 29 Palms - Flight 178-83 -- (Imbiorski) 14 images
- 1986 29 Palms Valley - Flight C461 -- (SBCFCD) 127 images Index
- 1990 Johnson - Morongo - Flight C483 -- (SBCFCD) 220 images
- 1993 Yucca Mesa - Flight C507 -- (SBCFCD) 4 images Index
- 1993 Yucca Valley - Flight C506 -- (SBCFCD) 18 images Index
- 1997 Wonder Valley - Flight C533 -- (SBCFCD) 281 images Index1 Index2
- 2003 Yucca Valley - Flight C547 -- (SBCFCD) 3 images
- 2008 29 Palms Flight 08-833 -- (Imbiorski) 3 images
-
Apple Valley (2,725 images)
Cities that are included in this section include Joshua Tree, Landers, Morongo, Pioneertown, Twenty-nine Palms, Wonder Valley, and Yucca Valley.
- 1933 Cleghorn Mountain - Flight 3678 -- (Rowe) 6 images
- 1933 Mojave River - Flight C2688 -- (Rowe) 7 images
- 1955 Mojave River - Flight F-36-2 -- (SBCFCD) 20 images Index
- 1955 Mojave River - Flight F-36-3 -- (SBCFCD) 17 images Index
- 1955 Mojave River - Flight F-36-4 -- (SBCFCD) 69 images Index
- 1960 Bear Valley Road - Flight C28 -- (SBCFCD) 18 images Index
- 1961 Apple Valley - Flight C23 -- (SBCFCD) 52 images Index
- 1962 Mojave River - Flight C20 -- (SBCFCD) 11 images Index
- 1962 Mojave River West Fork - Flight C18 -- (SBCFCD) 30 images Index
- 1963 Helendale - Flight C21 -- (SBCFCD) 29 images Index
- 1963 Victorville - Flight C19 -- (SBCFCD) 22 images Index
- 1964 East Desert Bowden Flat - Flight E-40 -- (SBCFCD) 108 images Index
- 1964 East Desert Landers - Flight E-39 -- (SBCFCD) 108 images Index
- 1964 East Desert Moronga - Flight E-41 -- (SBCFCD) 113 Index
- 1964 East Desert Morongo Valley - Flight E-42 -- (SBCFCD) 97 images Index
- 1964 Lucerne Valley - Flight C10 -- (SBCFCD) 30 images Index
- 1964 Oro Grande - Victorville - Flight C11 -- (SBCFCD) 14 images Index
- 1965 Oro Grande - Flight C98 -- (SBCFCD) 9 images Index
- 1965 Oro Grande - Flight C99 -- (SBCFCD) 13 images Index
- 1966 Air Base Road / Victorville - Flight C150 -- (SBCFCD) 36 images Index
- 1966 Air Base Road / Adelanto - Flight C151 -- (SBCFCD) 5 images Index
- 1966 Old Woman Springs - Flight C142 -- (SBCFCD) 24 images Index
- 1966 Stoddard Wells Road - Flight C155 -- (SBCFCD) 17 images Index
- 1967 Harper Lake Road - Flight C119 -- (SBCFCD) 8 images Index
- 1967 Harper Lake Road - Flight C120 -- (SBCFCD) 5 images Index
- 1970 Apple Valley - Flight C175 -- (SBCFCD) 10 images Index
- 1972 Apple Valley - Flight C207 -- (SBCFCD) 3 images
- 1972 Lucerne Valley - Flight C196 -- (SBCFCD) 30 images Index
- 1972 Mojave River - Flight C197 -- (SBCFCD) 19 images Index
- 1972 Victorville - Flight C192 -- (SBCFCD) 6 images
- 1973 Bell Mountain Road - Flight C236 -- (SBCFCD) 16 images
- 1973 Hesperia - Flight C211 -- (SBCFCD) 6 images
- 1973 Oro Grande Wash - Flight C202 -- (SBCFCD) 23 images Index
- 1974 Lucerne Valley - Flight C246 -- (SBCFCD) 24 images Index
- 1974 Victorville Exchange - Flight C253 -- (SBCFCD) 249 images Index
- 1975 Adelanto East Channel - Flight C254 -- (SBCFCD) 9 images Index
- 1975 Desert Knolls - Flight C259 -- (SBCFCD) 8 images
- 1975 Desert Knolls Wash - Flight C265 -- (SBCFCD) 9 images
- 1975 Mojave River - Flight C261 -- (SBCFCD) 64 images
- 1976 Lucerne Dry Lake - Flight C326 -- (SBCFCD) 27 images Index
- 1977 Lucerne Valley - Flight C313 -- (SBCFCD) 2 images Index
- 1977 Lucerne Valley - Flight C281 -- (SBCFCD) 20 images Index
- 1978 Mojave River - Deep Creek - Flight C298 -- (SBCFCD) 30 images Index
- 1979 Apple Valley - Hesperia - Flight C292 -- (SBCFCD) 35 images Index
- 1979 Hesperia - Flight C306 -- (SBCFCD) 11 images Index
- 1979 Mojave River - Flight C303 -- (SBCFCD) 29 images Index
- 1979 Mojave River - Hesperia - Flight C301 -- (SBCFCD) 19 images Index
- 1983 Apple Valley-Victorville - Flight C435 -- (SBCFCD) 18 images Index
- 1983 Apple Valley - Flight C436-1 -- (SBCFCD) 252 images Index
- 1983 Apple Valley (cont.) - Flight C436-2 -- (SBCFCD) 60 images Index
- 1983 Mojave River - Flight C433 -- (SBCFCD) 150 images IndexA IndexB
- 1984 Mojave Dam - Flight C445 -- (SBCFCD) 2 images
- 1990 Lucerne - Flight C481 -- (SBCFCD) 116 images
- 1991 Desert Knolls - Hesperia - Apple Valley - Flight C484 -- (SBCFCD) 131 images
- 1993 Bear Valley Road - Flight C511 -- (SBCFCD) 11 images
- 1993 Hesperia - Flight C496 -- (SBCFCD) 5 images Index
- 1993 I-15 Highway - Flight 509 -- (SBCFCD) 2 images
- 1993 Mojave River - Flight C497 -- (SBCFCD) 117 images IndexA IndexB
- 1993 Mojave River - Flight C504 -- (SBCFCD) 122 images Index1 Index2 Index3 Index4
- 1993 Seneca - Flight C510 -- (SBCFCD) 3 images
- 1993 Yates - Flight C508 -- (SBCFCD) 4 images
- 1994 Mojave River - Flight C517 -- (SBCFCD) 28 images Index
- 1994 Mojave River - Flight C518 -- (SBCFCD) 29 images Index
- 1995 Mojave River - Flight C519 -- (SBCFCD) 52 images Index
- 1995 Mojave River - Flight C520 -- (SBCFCD) 43 images Index
- 1997 Hesperia Basin - Flight C530 -- (SBCFCD) 3 images
- 1998 Oro Grande Basin - Flight 529 -- (SBCFCD) 4 images
- 2004 Hesperia - Flight C543 -- (SBCFCD) 20 images
-
Baker (191 images)
Cities that are included in this section include Baker, Cima, Ivanpah, Kelso, Kingston, Mountain Pass, and Nipton.
- 1966 Baker - Flight C250 -- (SBCFCD) 5 images
- 1973 Baker - Flight C208 -- (SBCFCD) 6 images
- 1977 Baker - Flight C315 -- (SBCFCD) 65 images Index Index
- 1978 Donnell - Flight C454 -- (SBCFCD) 2 images
- 1990 Kelbaker Road - Flight C485 -- (SBCFCD) 111 images
-
Baldy Mesa (917 images)
Cities that are included in this section include Adelanto, Baldy Mesa, El Mirage, Mountain View Acres, Oak Hills, Phelan, Pinion Hills, Silver Lakes, and Wrightwood.
- 1933 Mojave River, Landscape Images -- (Rowe) 7 images
- 1933 West Fork of the Mojave River Area - Flight 2688 -- (Rowe) 8 images
- 1936 Mojave River - Flight 3821 -- (Rowe) 4 images
- 1963 Mojave River - Flight C14 -- (SBCFCD) 21 images Index
- 1968 Phelan Road - Flight C168 -- (SBCFCD) 10 images Index
- 1969 Mojave River - Deep Creek - Flight C296 -- (SBCFCD) 58 images Index1 Index2
- 1972 Adelanto - Flight C206 -- (SBCFCD) 7 images
- 1972 Phelan - Flight C205 -- (SBCFCD) 6 images
- 1974 Wrightwood - Flight C260 -- (SBCFCD) 12 images
- 1974 Wrightwood - Flight C282 -- (SBCFCD) 12 images Index
- 1975 Adelanto East Channel - Flight C266 -- (SBCFCD) 7 images Index
- 1979 Wrightwood - Flight C290 -- (SBCFCD) 7 images Index
- 1982 Phelan - Flight 21-82 -- (Imbiorski) 2 images
- 1983 Adelanto - Flight C442-1 -- (SBCFCD) 252 images Index
- 1983 Adelanto (Cont) - Flight C442-2 -- (SBCFCD) 230 images Index
- 1991 Sheep Creek - Phelan - Flight C486 -- (SBCFCD) 8 images
- 1995 Mojave River - Flight C521 -- (SBCFCD) 51 images
- 2005 Mojave River - Flight C552 -- (SBCFCD) 206 images
-
Barstow (2,518 images)
Cities that are included in this section include Barstow, Barstow Heights, Calico, Daggett, Harvard, Hinckley, Lenwood, Ludlow, Newberry Springs, and Yermo.
- 1929 Mojave River Area - Flight 679 -- (Rowe) 164 images
- 1933 Mojave River - Flight 3685 -- (Rowe) 1 image
- 1939 Mojave River-Near Barstow -- (Rowe) 12 images
- 1939 San Gabriel River - Flight 3821 --(Rowe) 4 images
- 1960 Barstow - Flight C30 -- (SBCFCD) 74 images Index
- 1960 Lenwood - Flight C29 -- (SBCFCD) 34 images Index
- 1962 Cady Ranch-Mojave River - Flight C378 -- (SBCFCD) 8 images
- 1963 Hinkley Road - Flight C15 -- (SBCFCD) 21 images Index
- 1964 Mojave river - Flight C12 -- (SBCFCD) 42 images Index
- 1964 Toomey/Camp Cady - Flight C13 -- (SBCFCD) 28 images Index
- 1967 Jackhammer Gap - Flight C156 -- (SBCFCD) 6 images Index
- 1969 Mojave River - Yermo - Flight C294 -- (SBCFCD) 27 images Index
- 1970 Mojave River - Flight C179-- (SBCFCD) 27 images
- 1972 Barstow - Flight C209 -- (SBCFCD) 6 images Index
- 1972 Daggett - Flight C210 -- (SBCFCD) 6 images Index
- 1972 Kramer - Flight C213 -- (SBCFCD) 3 images
- 1972 Lenwood / Hinckley- Flight C214 -- (SBCFCD) 6 images Index
- 1972 Yermo - Flight C221 -- (SBCFCD) 6 images Index
- 1973 Newberry - Flight C217 -- (SBCFCD) 6 images Index
- 1978 Mojave River - Flight 304 -- (SBCFCD) 23 images Index
- 1978 Mojave River - Flight C305 -- (SBCFCD) 28 images Index
- 1979 Barstow-Daggett - Flight C307 -- (SBCFCD) 6 images Index
- 1979 Mojave River - Flight C302 -- (SBCFCD) 28 images Index
- 1979 West Barstow - Flight C308 -- (SBCFCD) 6 images Index
- 1980 Barstow - Flight C318 -- (SBCFCD) 21 images Index
- 1980 Lenwood Spreading Grounds - Flight C350 -- (SBCFCD) 2 images
- 1984 Barstow - Flight C443 -- (SBCFCD) 20 images
- 1984 Daggett - Newberry - Flight 444#1 -- (SBCFCD) 251 images Index
- 1984 Daggett - Newberry - Flight 444#2 -- (SBCFCD) 79 images Index
- 1989 East Desert - Flight C477 -- (SBCFCD) 144 images
- 1989 Newberry - Flight C475 -- (SBCFCD) 55 images
- 1989 North Desert - Flight C473(#1) -- (SBCFCD) 252 images
- 1989 North Desert - Flight C473(#2) -- (SBCFCD) 247 images
- 1990 Newberry - Flight C482 -- (SBCFCD) 126 images
- 1992 Mojave River - Flight C489 -- (SBCFCD) 67 images
- 1992 Mojave River - Flight C490 -- (SBCFCD) 86 images IndexA1 IndexA2 Index B1 IndexB2
- 1993 Calico - Flight C494 -- (SBCFCD) 5 images Index
- 1993 Mojave River - Flight C498 -- (SBCFCD) 71 images Index1 Index2 Index3 Index4
- 1993 Mojave River - Flight C505 -- (SBCFCD) 78 images Index1 Index2 Index3 Index4
- 1993 Newberry Springs - Flight C495 -- (SBCFCD) 11 images
- 1994 Mojave River - Flight C513 -- (SBCFCD) 15 images Index
- 1995 Mojave River - Flight C522 -- (SBCFCD) 131 images
- 2010 Mojave River - Flight C555 -- (SBCFCD) 125 images
- Barstow -- undated -- (Rowe) 131 images
- Barstow - Daggett -- (Imbiorski) 1 image
- 1985 Ludlow to Essex Road - Flight CT-8511-46 -- (CalTrans) 43 images
- 2004 Barstow to I-40 - Flight CT-0413-20A -- (CalTrans) 10 images
-
Big Bear (98 images)
Cities that are included in this section include Baldwin Lake, Bear City, Big Bear City, Big Bear Lake, Erwin Lake, Fawn Skin, Lake Williams, Moonridge, and Sugar Loaf.
- 1930 Baldwin Lake -- (Rowe) 12 images
- 1969 Bluff Lake - Flight C164 -- (SBCFCD) 2 images
- 1972 Big Bear - Flight C199 -- (SBCFCD) 8 images Index
- 1973 Big Bear - Flight C224 -- (SBCFCD) 15 images Index
- 1979 Big Bear - Flight C287 -- (SBCFCD) 58 images Index
-
Big River (367 images)
Cities that are included in this section include Big River, Earp, Havasu Lake, Parker, and Vidal.
- 1928 Colorado River - Flight C300 -- (Rowe) 4 images
- 1939 Colorado River - Flight AXM-203 -- (Rowe) 4 images
- 1940 Colorado River - Flight AXJ-320 -- (Rowe) 2 images
- 1941-45 Colorado River - Flight 2-2512 -- (Rowe) 68 images
- 1944 Colorado River - Flight 8924 -- (Rowe) 99 images
- 1948 Colorado River - Flight 12720 -- (Rowe) 3 images
- 1949 Colorado River - Flight 14613 -- (Rowe) 20 images
- 1949 Colorado River - Flight 14615 -- (Rowe) 14 images
- 1952 Colorado River - Flight 17727 -- (Rowe) 4 images
- 1957 Colorado River - Flight 7-20-57 -- (Rowe) 37 images
- 1959 Tract 5962 - Lake Havasu - Flight C106 -- (SBCFCD) 10 images
- 1972 Parker - Flight C218 -- (SBCFCD) 3 images
- 1983 Colorado River-Parker - Flight C439 -- (SBCFCD) 44 images Index
- 1990 Parker - C554 -- (SBCFCD) 54 images
-
Blue Jay (431 images)
Cities that are included in this section include Arrowbear, Blue Jay, Cedar Glen, Fredalba, Grass Valley, Green Valley Lake, Lake Arrowhead, Running Springs, and Twin Peaks.
- 1929 City Creek - Flight C-300 -- (Rowe) 2 images
- 1929 San Bernardino Mountains - Flight C300 -- (Rowe) 1 image
- 1932 South side of San Bernardino Mountains - Flight 2386 -- (Rowe) 64 images
- 1937 South side of San Bernardino Mountains - Flight 4646 -- (Rowe) 14 images
- 1938 Bear River Norton - Flight 4950 -- (Rowe) 4 images
- 1939 City Creek - Flight C5526 -- (Rowe) 1 image
- 1957 Bear River Norton - Flight GS-VQB -- (Rowe) 8 images
- 1965 Northeast San Bernardino Valley - Flight C139 -- (SBCFCD) 60 images Index
- 1966 Lake Arrowhead - Flight C455 -- (SBCFCD) 3 images
- 1972 Lake Arrowhead - Flight C190 -- (SBCFCD) 36 images
- 1974 Green Valley Lake - Flight C247 -- (SBCFCD) 11 images
- 1979 Crestline to Green Valley - Flight C285 -- (SBCFCD) 53 images Index
- 1994 City Creek - Flight 6864 -- (Imbiorski) 66 images
- 2004 City Creek - Flight C551 -- (SBCFCD) 80 images
- 2004 Cedar Glen - Flight C544 -- (SBCFCD) 26 images
-
Chino (2,252 images)
Cities that are included in this section include Carbon Canyon, Chino, Chino Hills, Montclair, Ontario, Rancho Cucamonga, San Antonio Heights, and Upland.
- 1927 Carbon Canyon - Flight 113619 -- (Rowe) 1 image
- 1930 Carbon Canyon - Flight 883031 -- (Rowe) 2 images
- 1930 Cucamonga - Flight 829 -- (Rowe) 5 images
- 1930 Cucamonga - Flight 883 -- (Rowe) 1 image
- 1930 Cucamonga - Flight 2510 -- (Rowe) 15 images
- 1930 Prado Dam - Flight 144 -- (Rowe) 13 images
- 1931_Prado_Dam - Flight C1740A -- (Rowe) 3 images
- 1937 Carbon Canyon - Flight 37-5 -- (Rowe) 2 images
- 1938 Carbon Canyon - Flight 38-19 -- (Rowe) 9 images
- 1938 Cucamonga Creek - Flight C263 -- (SBCFCD) 2 images
- 1938 Cucamonga Creek - Flight 5042 -- (Rowe) 9 images
- 1939 Chino-Prado Dam - Flight 5928 -- (Rowe) 62 images
- 1942 Carbon Canyon - Flight RSM -- (Rowe) 3 images
- 1945 Carbon Canyon -- (Rowe) 4 images
- 1947 Carbon Canyon -- Flight 11351 -- (Rowe) 1 image
- 1947 Prado Dam area -- (Rowe) 7 images
- 1948 Prado Basin - Flight C90 -- (SBCFCD) 29 images
- 1948 Prado Basin - Flight C91 -- (SBCFCD) 29 images
- 1951 Carbon Canyon - Flight 2 -- (Rowe) 5 images
- 1952 Prado Basin - Flight F-33-1 -- (SBCFCD) 71 images Index
- 1952 Prado Basin - Flight F-33-2 -- (SBCFCD) 16 images Index
- 1952 Prado Dam-Santa Ana River - Flight 17684 -- (Rowe) 4 images
- 1952 Santa Ana River/Prado Basin - Flight F-38 -- (SBCFCD) 39 images
- 1955 Chino Basin - Flight C464 -- (SBCFCD) 1 image
- 1955 Chino Basin - Flight F-34-1 -- (SBCFCD) 64 images Index
- 1955 Chino Basin - Flight F-34-2 -- (SBCFCD) 65 images Index
- 1955 Chino Basin - Flight F-34-3 -- (SBCFCD) 34 images Index
- 1955 Chino Basin - Flight F-34-4 -- (SBCFCD) 86 images Index
- 1955 Chino Basin - Flight F-34-5 -- (SBCFCD) 34 images
- 1955 Chino Basin - Flight F-34-6 -- (SBCFCD) 34 images
- 1955 Chino Basin - Flight F-34-7 -- (SBCFCD) 57 images
- 1955 Chino Basin - Flight F-34-8 -- (SBCFCD) 10 images
- 1955 Chino Basin - Flight F-34-9 -- (SBCFCD) 17 images
- 1960 Deer Creek - Flight C24 -- (SBCFCD) 43 images Index
- 1960 Prado Dam -- (Rowe) 1 image
- 1961 Cucamonga Storm Drain - Flight C17--(SBCFCD) 17 Index
- 1963 Carbon Canyon - Flight C133 -- (SBCFCD) 7 images Index
- 1963 Cucamonga Creek - Flight C1 -- (SBCFCD) 16 images Index
- 1963 Cucamonga Creek - Flight C2 -- (SBCFCD) 15 images Index
- 1963 Cucamonga Wash - Flight C145 -- (SBCFCD) 23 images Index
- 1964 Cucamonga Creek - Flight C5 -- (SBCFCD) 7 images Index
- 1964 Cucamonga Creek - Flight C6 -- (SBCFCD) 14 images Index
- 1965 Cucamonga Wash - Flight C100 -- (SBCFCD) 44 images Index
- 1965 Cucamonga Wash - Flight C101 -- (SBCFCD) 58 images Index
- 1965 Deer Creek - Flight C147 -- (SBCFCD) 2 images
- 1966 Deer - Day Channel - Flight C163 -- (SBCFCD) 7 images
- 1966 Deer - Day Creeks - Flight C105 -- (SBCFCD) 18 images Index
- 1966 San Sevaine - Flight C109 -- (SBCFCD) 18 images Index
- 1967 Cypress Channel - Flight C118 -- (SBCFCD) 6 images Index
- 1968 Cucamonga Creek - Flight C264 -- (SBCFCD) 2 images Index
- 1970 Cucamonga Creek - Flight C262 -- (SBCFCD) 8 images Index
- 1970 Mt. Baldy - Flight C178 -- (SBCFCD) 6 images
- 1971 Cucamonga Creek - Flight C262A -- (SBCFCD) 4 images
- 1972 Prado Basin - Flight C195 -- (SBCFCD) 13 images Index
- 1972 West San Bernardino Valley - Flight C193 -- (SBCFCD) 42 images Index
- 1973 Milliken Disposal - Flight C257 -- (SBCFCD) 6 images Index
- 1973 San Sevaine Channel - Flight C379 -- (SBCFCD) 4 images
- 1974 Alta Loma Basin - Flight C240 -- (SBCFCD) 7 images Index
- 1974 Day Creek Canyon - Flight C239 -- (SBCFCD) 32 images Index
- 1974 Montclair Basin - Flight C244 -- (SBCFCD) 3 images Index
- 1974 Montclair Basin - Flight C248 -- (SBCFCD) 4 images Index
- 1975 Chino Creek - Flight C371 -- (SBCFCD) 7 images
- 1975 Chino Hills - Flight C252 -- (SBCFCD) 82 images Index
- 1975 Day Creek Spreading Ground - Flight C267 -- (SBCFCD) 13 images Index
- 1976 Cucamonga Creek - Flight C274 -- (SBCFCD) 11 images Index
- 1978 Alta Loma-197-78 -- (Imbiorski) 14 images
- 1978 Cable Creek Channel - Flight C370A -- (SBCFCD) 1 image
- 1978 Cucamonga Channel - Flight C370B -- (SBCFCD) 3 images
- 1978 Cucamonga Creek - Flight C322 -- (SBCFCD) 90 images Index Index Index
- 1978 Day Creek Channel - Flight C310 -- (SBCFCD) 15 images Index
- 1978 Day Creek Channel - Flight C370C -- (SBCFCD) 4 images
- 1978 Deer Creek Channel - Flight C370D -- (SBCFCD) 5 images
- 1978 Dunlop Channel - Flight C370E -- (SBCFCD) 1 image
- 1978 Etiwanda Creek Channel - Flight C370F -- (SBCFCD) 3 images
- 1978 Lytle-Cajon - Flight C370I -- (SBCFCD) 1 image
- 1978 Ontario - Flight 7859 -- (Imbiorski) 29 images
- 1978 San Bernardino Valley Oblique - Flight C370 -- (SBCFCD) 6 images Index
- 1978 San Sevaine - Flight C312 -- (SBCFCD) 1 image Index
- 1978 San Sevaine Channel - Flight C370G -- (SBCFCD) 7 images
- 1978 San Timoteo Channel - Flight C370H -- (SBCFCD) 4 images
- 1978 West Cucamonga Channel - Flight C299 -- (SBCFCD) 12 images
- 1978 West Cucamonga Channel - Flight C280 -- (SBCFCD) 82 images
- 1978 West Cucamonga Creek - Flight C309 -- (SBCFCD) 26 images Index
- 1979 15th Street Interceptor - Flight C363 -- (SBCFCD) 2 images Index
- 1979 College Heights - Flight 361 -- (SBCFCD) 2 images Index
- 1979 College Heights - Flight 362 -- (SBCFCD) 5 images Index
- 1979 Cucamonga Basin - Flight C459 -- (SBCFCD) 5 images
- 1979 Deer Creek Bridges - Flight C340 -- (SBCFCD) 40 images Index
- 1979 Demens Creek - Flight C323 -- (SBCFCD) 8 images Index
- 1979 San Antonio Basins - Flight C356 -- (SBCFCD) 1 image Index
- 1980 Chino- Flight 129-80 -- (Imbiorski) 5 images
- 1980 Chino - Flight 229-80 -- (Imbiorski) 2 images
- 1980 Cucamonga-Guasti Park - Flight C321 -- (SBCFCD) 14 images Index
- 1980 Deer Creek - Flight C376 -- (SBCFCD) 8 images
- 1980 Prado Dam - Flight C320 -- (SBCFCD) 7 images Index
- 1980 Prado Dam - Flight C341 -- (SBCFCD) 4 images
- 1980 San Sevaine - Flight C343 -- (SBCFCD) 6 images
- 1981 Chino - Flight 282-81 -- (Imbiorski) 4 images
- 1981 Cucamonga Basin - Flight C377 -- (SBCFCD) 4 images
- 1981 Cucamonga Creek - Flight C385 -- (SBCFCD) 2 images
- 1981 Ontario - Flight 309-81 -- (Imbiorski) 5 images
- 1981 Rancho Cucamonga - Flight 110-81 -- (Imbiorski) 3 images
- 1981 Rancho Cucamonga - Flight 79-81 -- (Imbiorski) 12 images
- 1981 Rancho Cucamonga - Flight 151-81 -- (Imbiorski) 2 images
- 1981 San Antonio Heights - Flight C440 -- (SBCFCD) 12 images
- 1981 San Anotnio Heights Oblique - Flight C441 -- (SBCFCD) 2 images
- 1981 San Sevaine - Flight C342 -- (SBCFCD) 9 images Index
- 1981 Upland - Flight 113-81 -- (Imbiorski) 2 images
- 1981 Upland - Flight 116-81 -- (Imbiorski) 9 images
- 1981 Upland - Flight 230-81 -- (Imbiorski) 2 images
- 1982 Alta Loma - Flight 189-82 -- (Imbiorski) 40 images
- 1982 Day Creek - Flight C426 -- (SBCFCD) 7 images
- 1982 Rancho Cucamonga - Flight 148-82 -- (Imbiorski) 3 images
- 1983 Chino - Flight C432 -- (SBCFCD) 87 images
- 1983 Day Creek Spreading Ground - Flight C434 -- (SBCFCD) 7 images
- 1983 Los Serranos - Flight C431 -- (SBCFCD) 6 images Index
- 1983 Rancho Cucamonga - Flight 76-83 -- (Imbiorski) 3 images
- 1983 Rancho Cucamonga - Flight 110-83 -- (Imbiorski) 2 images
- 1984 Chino - Flight 88-84 -- (Imbiorski) 10 images
- 1984 Rancho Cucamonga - Flight 58-84 -- (Imbiorski) 2 images
- 1984 Upland - Flight 136-84 -- (Imbiorski) 6 images
- 1985 Chino Airport - Flight C456 -- (SBCFCD) 13 images
- 1985 Chino Airport - Flight C457 -- (SBCFCD) 12 images
- 1985 Cucamonga Creek - Flight C446 -- (SBCFCD) 12 images
- 1985 Upland - Flight 165-85 -- (Imbiorski) 4 images
- 1986 Montclair - Flight C476 -- (SBCFCD) 4 images
- 1986 Ontario - Flight C453 -- (SBCFCD) 2 images
- 1986 Rancho Cucamonga - Flight 176-86 -- (Imbiorski) 2 images
- 1986 Upland - Flight 73-86 -- (Imbiorski) 4 images
- 1987 Chino Flight 180-87 -- (Imbiorski) 2 images
- 1987 Montclair - Flight 157-87 -- (Imbiorski) 3 images
- 1987 Rancho Cucamonga - Flight 98-87 -- (Imbiorski) 2 images
- 1987 Rancho Cucamonga - Flight 151-87 -- (Imbiorski) 2 images
- 1987 Rancho Cucamonga - Flight 232-87 -- (Imbiorski) 2 images
- 1987 Upland - Flight 3-87 -- (Imbiorski) 9 images
- 1988 Chino - Flight 19-88 -- (Imbiorski) 3 images
- 1989 Rancho Cucamonga - Flight 287-89 -- (Imbiorski) 2 images
- 1989 Upland - Flight 116-89 -- (Imbiorski) 2 images
- 1990 Deer Creek - Flight C480 -- (SBCFCD) 3 images
- 1991 Chino -- (Imbiorski) 1 image
- 1991 Upland - Flight 150-91 -- (Imbiorski) 8 images
- 1992 Chino - Flight 24-92 -- (Imbiorski) 5 images
- 1993 Chino - Flight 128-93 -- (Imbiorski) 4 images
- 1993 Ontario - Flight 75-93 -- (Imbiorski) 2 images
- 1993 Rancho Cucamonga - Flight 34-93 -- (Imbiorski) 2 images
- 1993 San Sevaine - Flight C502 -- (SBCFCD) 7 images
- 1993 San Sevaine - Flight C512 -- (SBCFCD) 32 images Index
- 1994 Chino Flight 91-94 -- (Imbiorski) 10 images
- 1994 Chino Hills - Flight 6862 -- (NRCS) 8 images
- 1994 San Antonio Creek Canyon - Flight 6851 -- (NRCS) 9 images
- 1994 Santa Ana River - Flight C516 -- (SBCFCD) 5 images
- 1996 Chino -- (Imbiorski) 1 image
- 1997 Cucamonga Channel - Flight C535 -- (SBCFCD) 6 images
- 1997 Ontario - Flight 31-97 -- (Imbiorski) 4 images
- 1997 San Antonio Creek - Flight C534 -- (SBCFCD) 22 images
- 1998 Ontario - Flight 38-98 -- (Imbiorski) 2 images
- 2000 Rancho Cucamonga Flight 849-00 -- IImbiorski) 27 images
- 2000 Rancho Cucamonga - Flight C539 -- (SBCFCD) 10 images
- 2003 Upland - Flight 15-03 -- (Imbiorski) 2 images
- 2004 Cucamonga Creek - Flight C549 -- (SBCFCD) 10 images
- 2007 Rancho Cucamonga - Flight 843-07 -- (Imbiorski) 4 images
- No date - Deer - Day Creek - Flight C458 -- (SBCFCD) 4 images
-
Crestline (989 images)
Cities that are included in this section include Cedar Pines Park, Crestforest, and Crestline.
- 1933 Valley of the Moon - Flight 2688 -- (Rowe) 4 images
- 1939 Rim of the World - Flight 6090 -- (Rowe) 23 images
- 1958 Waterman Canyon -- (Rowe) 4 images
- 1960 Waterman Canyon -- (Rowe) 5 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 1 -- (USDA-SBCFCD) 92 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 2 -- (USDA-SBCFCD) 72 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 3-1 -- (USDA-SBCFCD) 247 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 3-2 -- (USDA-SBCFCD) 9 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 4 -- (USDA-SBCFCD) 76 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 5 -- (USDA-SBCFCD) 167 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 6 -- (USDA-SBCFCD) 12 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 8 -- (USDA-SBCFCD) 65 images
- 1965 National Forest-Big Bear to Mt. Baldy - Flight EPH 9 -- (USDA-SBCFCD) 71 images
- 1992 Crest Forest - Flight C492 -- (SBCFCD) 85 images Index
- 1995 Rim Forest - Flight C524 -- SBCFCD) 12 images Index
- 1995 Rim Forest - Flight C527 -- (SBCFCD) 12 images
- 2004 Waterman Canyon - Flight C550 -- (SBCFCD) 31 images
-
Fontana (1,363 images)
Cities that are included in this section include Bloomington, Cajon, Crestmore, Devore, Etiwanda, Fontana, Keenbrook, Lytle Creek, and Rialto.
- 1930 Cajon Pass - Flight 910 -- (Rowe) 3 images
- 1930 Ontario-Etiwanda area - Flight 829 -- (Rowe) 64 images
- 1930 San Gabriel Mountains - Flight 819 -- (Rowe) 26 images
- 1932 Cajon Pass - Flight 2388 -- (Rowe) 17 images Index
- 1932 Cajon Pass - Flight 2387 -- (Rowe) 16 images
- 1933 Cajon Pass - Flight 2510 -- (Rowe) 24 images
- 1933 Cajon Pass - Flight 2580 -- (Rowe) 3 images
- 1936 Cajon Pass - Flight G -- (Rowe) 7 images
- 1938 Cajon Pass - Flight 5029 -- (Rowe) 2 images
- 1943 Cajon Pass -- Flight 8236 -- (Rowe) 47 images
- 1943 Cajon Pass – Flight 8305 -- (Rowe) 3 images
- 1943 Lytle Creek - Flight 8305 -- (Rowe) 40 images
- 1945 Cajon Pass -- Flight BC-2 -- (Rowe) 3 images
- 1961 Rialto Storm Drain - Flight C25 -- (SBCFCD) 12 images Index
- 1962 Lytle Creek - Flight C16 -- (SBCFCD) 18 Index
- 1963 Fontana - Flight C7 -- (SBCFCD) 31 images Index
- 1964 Lytle Creek - Flight C141 -- (SBCFCD) 24 images Index
- 1964 Rialto - Flight C3 -- (SBCFCD) 12 images Index
- 1964 Rialto - Flight C4 -- (SBCFCD) 20 images Index
- 1965 Northwest San Bernardino Valley - Flight C140 -- (SBCFCD) 45 images Index
- 1966 East Etiwanda & San Sevaine - Flight C104 -- (SBCFCD) 25 images Index
- 1966 East Fontana - Flight C103 -- (SBCFCD) 10 images Index
- 1966 Lytle Creek - Flight C144 -- (SBCFCD) 26 images
- 1966 Lytle Creek - Flight C146 -- (SBCFCD) 3 images
- 1966 Sierra Ave. - Stover Road - Flight C115 -- (SBCFCD) 25 images
- 1967 Lytle Creek - Flight C117 -- (SBCFCD) 3 images
- 1967 Lytle Creek - Flight C132 -- (SBCFCD) 24 images Index
- 1967 Lytle Creek - Flight C136 -- (SBCFCD) 10 images Index
- 1968 Cactus Basin - Flight C154 -- (SBCFCD) 9 images Index
- 1970 San Bernardino County Flight -Flight 11-24-1970 -- (SBCA) 119 images
- 1971 Cucamonga Creek - Flight C187 -- (SBCFCD) 1 image
- 1972 Devore - Flight C237 -- (SBCFCD) 5 images
- 1972 Lytle Creek - Flight C182 -- (SBCFCD) 14 images Index
- 1973 Fontana/Bloomington - Flight C241 -- (SBCFCD) 14 images
- 1973 Rich Basin - Flight C200 -- (SBCFCD) 3 images
- 1974 Bloomington Avenue Drainage - Flight C255 -- (SBCFCD) 14 images
- 1974 Cajon Disposal Site - Flight C245 -- (SBCFCD) 2 images
- 1978 Daly Channel - Flight 324 -- (SBCFCD) 8 images Index
- 1979 Lytle Creek - Flight C286 -- (SBCFCD) 7 images Index
- 1980 San Sevaine - Flight C398 -- (SBCFCD) 9 images
- 1981 San Sevaine Basin - Flight C399 -- (SBCFCD) 9 images
- 1984 Fontana - Flight 141-84 -- (Imbiorski) 2 images
- 1984 Fontana - Flight 194-84 -- (Imbiorski) 1 image
- 1986 Fontana - Flight C460 -- (SBCFCD) 1 image
- 1988 Flood Basins Texas Fire - Flight C470 -- (SBCFCD) 32 images Index
- 1990 San Sevaine - Flight C478 -- (SBCFCD) 4 images
- 1990 San Sevaine - Flight C479 -- (SBCFCD) 15 images
- 1991 San Bernardino-Rialto - Flight 147-91 -- (Imbiorski) 2 images
- 1993 Devore - Flight 73-93 - (Imbiorski) 5 images
- 1994 Cajon Creek - Flight C514 -- (SBCFCD) 9 images Index
- 1994 Etiwanda - Flight 6852 -- (NRCS) 30 images
- 1994 Fontana - Flight 6866 -- (NRCS) 71 images
- 1996 Etiwanda - Flight 129-96 -- (Imbiorski) 4 images
- 1998 Cajon Creek Wash- Flight C536 -- (SBCFCD) 22 images Index
- 1999 Cajon Creek Wash - Flight C538 -- (SBCFCD) 33 images
- 1999 Lytle Creek - Flight 99-860 -- (Imbiorski) 9 images
- 1999 Lytle Creek - Flight 8602 -- (Imbiorski) 9 images
- 2004 Devore - Flight C545 -- (SBCFCD) 57 images
- 2004 Lytle Creek - Flight C546 -- (SBCFCD) 281 images
-
Needles (496 images)
Cities that are included in this section include Amboy, Essex, Geoffs, Needles, and Rice.
- 1928 Lower Colorado River near Needles -- (Rowe) 4 images
- 1930 Colorado River - Flight 911 -- (Rowe) 1 image
- 1933 Colorado River - Flight 2689 -- (Rowe) 1 image
- 1939 Lower Colorado Flight C-5750 -- (Rowe) 4 images
- 1940 Lower Colorado River - Flight AXJ-320 -- (Rowe) 2 images
- 1948 Lower Colorado - Flight 12720 -- (Rowe) 3 images
- 1949 Davis to Topock - Flight 14613 -- (Rowe) 21 images
- 1949 Lower Colorado River - Flight 14615 --(Rowe) 14 images
- 1952 Lower Colorado River - Flight 17727 -- (Rowe) 4 images
- 1953 Mohave Valley -- (Rowe) 8 images
- 1957 Colorado River - near Needles -- (Rowe) 37 images
- 1963 Needles - Flight C22 -- (SBCFCD) 21 images Index
- 1964 Ft. Mojave to Lake Havasu - Flight C122 -- (SBCFCD) 46 images Index
- 1967 Eagle Pass Wash - Flight C131 -- (SBCFCD) 3 images Index
- 1972 Needles - Flight C216 -- (SBCFCD) 3 images
- 1973 Needles - Flight C317 -- (SBCFCD) 6 images Index
- 1974 Needles - Flight C251 -- (SBCFCD) 2 images Index
- 1976 Eagle Pass - Flight C314 -- (SBCFCD) 3 images Index
- 1976 Needles - Flight C283 -- (SBCFCD) 91 images Index
- 1992 Needles - Flight C491 -- (SBCFCD) 47 image Index
- 2002 Needles - Flight 816 -- (Imbiorski) 97 images
- Colorado River - Flight 2-2512 -- (Rowe) 68 images
- Lower Colorado River Index Maps -- (Rowe) 2 images
- 2004 Barstow at I-40 - Flight CT-9412-48 -- (CalTrans) 31 images
-
San Bernardino East (1,334 images)
Cities that are included in this section include Angelus Oaks, Barton Flats, Calimesa, Crafton, Forest Falls, Forest Home, Jenks Lake, Mountain Home Village, Oak Glen, Seven Oaks, and Yucaipa.
- 1928 San Andreas Fault -- (Rowe) 1 image
- 1929 San Andreas Fault -- (Rowe) 1 image
- 1930 Mt. Home Creek - Flight 911 -- (Rowe) 9 images
- 1930 San Andreas Fault - Yucaipa to Oak Glen -- Flight 910 -- (Rowe) 15 images
- 1930 San Bernardino Mountains - Flight C910 -- (Rowe) 57 images
- 1931 San Andreas Fault - Flight 1740 -- (Rowe) 1 image
- 1948 Yucaipa Middle Bench - Flight U-37 -- (SBCFCD) 27 images Index
- 1955 East Valley - Flight F-35-1 -- (SBCFCD) 20 images Index
- 1955 East Valley - Flight F-35-2 -- (SBCFCD) 20 images Index
- 1955 East Valley - Flight F-35-3 -- (SBCFCD) 22 images Index
- 1955 East Valley - Flight F-35-4 -- (SBCFCD) 39 images Index
- 1955 East Valley - Flight F-35-5 -- (SBCFCD) 6 images
- 1955 East Valley - Flight F-35-6 -- (SBCFCD) 8 images
- 1955 East Valley - Flight F-35-7 -- (SBCFCD) 22 images
- 1955 East Valley - Flight F-35-8 -- (SBCFCD) 20 images
- 1955 East Valley - Flight F-35-10 -- (SBCFCD) 14 images
- 1957 Yucaipa - Flight C143 -- (SBCFCD) 4 images Index
- 1957 Yucaipa - Flight C223 -- (SBCFCD) 14 images
- 1963 Oak Glen Creek - Flight C27 -- (SBCFCD) 10 images Index
- 1963 Yucaipa - Flight C111 -- (SBCFCD) 11 images Index
- 1964 Live Oak Canyon - Flight C112 -- (SBCFCD) 21 images Index
- 1965 San Bernardino Valley East Landuse - Flight C137 - (SBCFCD) 45 images Index
- 1965 Sand Canyon - C162 -- (SBCFCD) 21 images Index
- 1965 Upper Zanja - Flight C102 -- (SBCFCD) 8 images Index
- 1965 Sixth Street, Yucaipa - Flight C110 -- (SBCFCD) 5 images Index
- 1966 Mill Creek - Flight C128 -- (SBCFCD) 17 images Index
- 1967 City Creek - Flight C148 -- (SBCFCD) 11 images Index
- 1967 Mill Creek - Flight C129 -- (SBCFCD) 5 images
- 1967 Mill & Zanja Creeks - Flight C130 -- (SBCFCD) 20 images Index
- 1967 Yucaipa - (Rowe) 44 images
- 1968 Wilson Creek / Wildwood Creek - Flight C149 -- (SBCFCD) 17 images Index
- 1969 Mill Creek - Flight C157 -- (SBCFCD) 17 images Index
- 1970 San Bernardino County Flight -Flight 1-2-1970 -- (SBCA) 81 images
- 1970 San Bernardino County Flight -Flight 1-30-1970 -- (SBCA) 73 images
- 1971 Sand Canyon - Flight C188 -- (SBCFCD) 12 images Index
- 1971 Wilson Creek / Wildwood Creek - Flight C184 -- (SBCFCD) 13 images Index
- 1971 Yucaipa Creek - Flight C185 -- (SBCFCD) 3 images
- 1972 East San Bernardino Valley - Flight C194 -- (SBCFCD) 31 images Index
- 1972 West Avenue / Yucaipa Creek- Flight C189 -- (SBCFCD) 10 images
- 1972 Yucaipa - Flight C238 -- (SBCFCD) 6 images
- 1974 Yucaipa - Flight C170 -- (SBCFCD) 3 images Index
- 1975 Yucaipa - Flight C273 -- (SBCFCD) 135 images Index 1 Index 2
- 1976 Mission Zanja Oblique - Flight C375 -- (SBCFCD) 4 images
- 1979 Forest Falls - Oak Glen - Flight C289 -- (SBCFCD) 23 images Index
- 1979 Wilson Basins - Flight C359 -- (SBCFCD) 2 images
- 1985 East Valley - Flight C447 -- (SBCFCD) 12 images
- 1987 Live Oak Canyon Road - Flight C462 -- (SBCFCD) 10 images Index
- 1987 Yucaipa Oblique - Flight C471 -- (SBCFCD) 12 images
- 1988 San Bernardino to Seven Oaks Dam - Flight IE-94 -- (Schnepp) 121 images Index
- 1988 Southeast of San Bernardino Valley - Flight C472 -- (SBCFCD) 26 images Index
- 1990 Plunge Creek - Flight C515 -- (SBCFCD) 6 images
- 1992 Forest Falls - Flight C493 -- (SBCFCD) 11 images Index
- 1993 Santa Ana River - Flight C503 -- (SBCFCD) 15 images Index
- 1993 Seven Oaks Dam - Flight C499 -- (SBCFCD) 5 images Index
- 1993 Seven Oaks Dam - Flight C500 -- (SBCFCD) 2 images
- 1994 Yucaipa - Flight 6877 -- (NRCS) 23 images
- 1995 Seven Oaks Dam - Flight C525 -- (SBCFCD) 9 images Index
- 1997 Mill Creek - Flight C531 -- (SBCFCD) 2 images
- 2000 Forest Falls - Flight C540 -- (SBCFCD) 4 images
- 2004 Yucaipa - Flight C556 -- (SBCFCD) 75 images
- 2004 Yucaipa - Flight C557 -- (SBCFCD) 14 images
-
San Bernardino West (5,240 images)
- 1930 San Bernardino Valley - Flight -- (Rowe) 128 images
- 1930 San Bernardino County - Flight C910 -- (Rowe) 59 images
- 1933 North Fork - Flight 2688 -- (Rowe) 7 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ34 -- (USDA-SBCFCD) 10 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ40 -- (USDA-SBCFCD) 10 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ41 -- (USDA-SBCFCD) 7 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ42 -- (USDA-SBCFCD) 9 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ60 -- (USDA-SBCFCD) 4 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ66 -- (USDA-SBCFCD) 14 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ67 -- (USDA-SBCFCD) 3 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ80 -- (USDA-SBCFCD) 8 images
- 1938 Santa Ana River (Post-Flood) - Flight AXJ81 -- (USDA-SBCFCD) 18 images
- 1938 Santa Ana River (Post-Flood) - Flight AXK39 -- (USDA-SBCFCD) 4 images
- 1938 Santa Ana River (Post-Flood) - Flight AXK40 -- (USDA-SBCFCD) 2 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL39 -- (USDA-SBCFCD) 15 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL40 -- (USDA-SBCFCD) 20 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL41 -- (USDA-SBCFCD) 32 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL42 -- (USDA-SBCFCD) 31 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL60 -- (USDA-SBCFCD) 42 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL61 -- (USDA-SBCFCD) 36 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL62 -- (USDA-SBCFCD) 20 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL63 -- (USDA-SBCFCD) 31 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL65 -- (USDA-SBCFCD) 28 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL67 -- (USDA-SBCFCD) 24 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL68 -- (USDA-SBCFCD) 22 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL73 -- (USDA-SBCFCD) 39 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL74 -- (USDA-SBCFCD) 45 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL76 -- (USDA-SBCFCD) 24 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL77 -- (USDA-SBCFCD) 29 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL78 -- (USDA-SBCFCD) 24 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL79 -- (USDA-SBCFCD) 91 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL80 -- (USDA-SBCFCD) 27 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL83 -- (USDA-SBCFCD) 56 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL84 -- (USDA-SBCFCD) 67 images
- 1938 Santa Ana River (Post-Flood) - Flight AXL85 -- (USDA-SBCFCD) 44 images
- 1938 Santa Ana River (Post-Flood) - Flight AXl102 -- (USDA-SBCFCD) 45 images
- 1938 Santa Ana River (Post-Flood) - Flight AXM39 -- (USDA-SBCFCD) 2 images
- 1938 Santa Ana River (Post-Flood) - Flight AXM40 -- (USDA-SBCFCD) 1 images
- 1938 Santa Ana River (Post-Flood) - Flight AXM41 -- (USDA-SBCFCD) 4 images
- 1938 Santa Ana River (Post-Flood) - Flight AXM61 -- (USDA-SBCFCD) 6 images
- 1938 Santa Ana River (Post-Flood) - Flight AXM62 -- (USDA-SBCFCD) 5 images
- 1938 Santa Ana River (Post-Flood) - Flight AXM73 -- (USDA-SBCFCD) 3 images
- 1938 Santa Ana River (Post-Flood) - Flight AXM74 -- (USDA-SBCFCD) 45 images
- 1938 San Bernardino City - Flight C383 -- (SBCFCD) 1 image
- 1939 East Highland - Flight 5526 -- (Rowe) 1 image
- 1943 Colton Area - Flight 8305 -- (Rowe) 4 images
- 1945 North Fork Canal - Flight 9800 -- (Rowe) 13 images
- 1960 Barton Road - San Timoteo Wash - Flight C116 -- (SBCFCD) 7 images
- 1962 San Bernardino & Colton - Flight C26 -- (SBCFCD) 28 images Index
- 1963 Piedmont Dr. - Muscoy - Flight C113 -- (SBCFCD) 7 images Index
- 1963 Reche Canyon - Flight C160 -- (SBCFCD) 10 images Index
- 1964 Piedmont Dr. - Arden - Flight C114 -- (SBCFCD) 10 images
- 1964 Piedmont Extension - Flight C169 -- (SBCFCD) 9 images Index
- 1964 Santa Ana River - Flight C9 -- (SBCFCD) 25 Index
- 1964 Santa Ana Wash - Flight C92 -- (SBCFCD) 28 images
- 1964 Tippecanoe Street - Flight C165 -- (SBCFCD) 23 images Index
- 1964 Upper Warm Creek - Flight C8 -- (SBCFCD) 46 images Index
- 1965 Southwest San Bernardino Valley - Flight C138 -- (SBCFCD) 41 images Index
- 1966 Baseline / Del Rosa - Flight C153 -- (SBCFCD) 5 images Index
- 1966 Cable Canyon - Flight C107 -- (SBCFCD) 22 images Index
- 1966 Cable Canyon - Flight C108 -- (SBCFCD) 17 images Index
- 1966 Upper Warm Creek - Flight C381 -- (SBCFCD) 1 image
- 1967 East San Bernardino Valley - Flight C183 -- (SBCFCD) 49 images Index
- 1969 San Bernardino Valley - Flight C295 -- (SBCFCD) 111 images
- 1969 West San Bernardino Valley - Flight C293 -- (SBCFCD) 88 images
- 1970 Barton Road - Flight C176 -- (SBCFCD) 17 images Index
- 1970 Mountain View - Flight C177 --(SBCFCD) 3 images
- 1971 San Bernardino Valley - Flight C186 -- (SBCFCD) 57 images Index
- 1973 Scott Canyon - Flight C225 -- (SBCFCD) 4 images Index
- 1974 Colton Disposal - Flight C256A -- (SBCFCD) 3 images Index
- 1977 San Bernardino Valley - Flight C2 -- (SBCFCD) 11 images
- 1978 San Bernardino Valley - Flight C279 -- (SBCFCD) 188 images Index 1 Index 2 Index 3
- 1978 Warm Creek - Flight 300 -- (SBCFCD) 2 images Index
- 1979 Daly Basin - Flight C347 -- (SBCFCD) 5 images
- 1979 Daly Basin - Flight C348 -- (SBCFCD) 3 images
- 1979 Harrison Basin - C364 -- (SBCFCD) 2 images
- 1979 Lemon Basin - Flight C357 -- (SBCFCD) 5 images
- 1979 Little Sand Dam - Flight C360 -- (SBCFCD) 2 images
- 1979 McQuiddy Basin - Flight C358 -- (SBCFCD) 7 images
- 1979 Patton Basin - Flight C346 -- (SBCFCD) 4 images
- 1979 Sand Basin - Flight C345 -- (SBCFCD) 2 images
- 1979 Sand Canyon - Flight C428 -- (SBCFCD) 5 images
- 1979 Small Canyon Basin - C354 -- (SBCFCD) 2 images
- 1980 Badger Canyon - Flight C423 -- (SBCFCD) 7 images
- 1980 Borea Canyon - Flight C327 -- (SBCFCD) 5 images
- 1980 Cook Canyon - FLight C336 -- (SBCFCD) 6 images
- 1980 Cook Canyon Basin - Flight C338 -- (SBCFCD) 2 images
- 1980 Daly Basin - Flight C349 -- (SBCFCD) 3 images
- 1980 Devil Canyon Basin - Flight C389 -- (SBCFCD) 3 images
- 1980 Devil Canyon - Flight C331 -- (SBCFCD) 4 images
- 1980 Dynamite Canyon Basin - Flight C391 -- (SBCFCD) 2 images
- 1980 Dynamite Canyon - Flight C332 -- (SBCFCD) 2 images
- 1980 Dynamite Canyon - Flight C339 -- (SBCFCD) 2 images
- 1980 East Twin Peak Line - Flight C328 -- (SBCFCD) 5 images
- 1980 East Twin Peak Line - Flight C329 -- (SBCFCD) 4 images
- 1980 Harrison Canyon - Flight C366 -- (SBCFCD) 7 images
- 1980 Harrison Canyon - Flight C367-1 -- (SBCFCD) 7 images
- 1980 Harrison Canyon - Flight C368 -- (SBCFCD) 29 images
- 1980 Harrison Canyon - Flight C369 -- (SBCFCD) 2 images
- 1980 Harrison Canyon - Flight C427 -- (SBCFCD) 6 images
- 1980 Harrison Canyon Oblique - Flight C367-2 -- (SBCFCD) 4 images
- 1980 Harrison Canyon - Flight C365 -- (SBCFCD) 7 images
- 1980 Lemon Basin - Flight C392 -- (SBCFCD) 3 images
- 1980 Little Mountain Dam - Flight C414 -- (SBCFCD) 4 images
- 1980 Little Mountain Dam - Flight C415 -- (SBCFCD) 4 images
- 1980 Little Sand Canyon - Flight C334 -- (SBCFCD) 8 images
- 1980 Little Sand Dam - Flight C344 -- (SBCFCD) 2 images
- 1980 Loma Linda - Flight C388 -- (SBCFCD) 8 images
- 1980 McQuiddy - Flight C394 -- (SBCFCD) 5 images
- 1980 McQuiddy-Brush - Flight C395 -- (SBCFCD) 5 images
- 1980 McQuiddy-Brush - Flight C396 -- (SBCFCD) 5 images
- 1980 McQuiddy-Brush - Flight C397 -- (SBCFCD) 5 images
- 1980 Mt. McKinley - Flight C374 -- (SBCFCD) 12 images
- 1980 Mt. Shadows Overflow - Flight C355 -- (SBCFCD) 1 image
- 1980 North & West Badger - Flight C422 -- (SBCFCD) 4 images
- 1980 Small Canyon - Flight C333 -- (SBCFCD) 5 images
- 1980 Small Canyon Basin - Flight C352 -- (SBCFCD) 2 images
- 1980 Small Canyon Basin - Flight C353 -- (SBCFCD) 2 images
- 1980 South Badger-Sycamore - Flight C417 -- (SBCFCD) 6 images
- 1980 South Badger Sycamore - Flight C418 -- (SBCFCD) 6 images
- 1980 Sycamore Canyon - Flight C420 -- (SBCFCD) 8 images
- 1980 Twin Peaks Spreading Ground - Flight C406 -- (SBCFCD) 6 images
- 1980 Waterman Basin - Flight C386 -- (SBCFCD) 2 images
- 1980 Waterman Canyon - Flight C330 -- (SBCFCD) 7 images
- 1980 Wiggins #2 - Flight C421 -- (SBCFCD) 5 images
- 1980 Wiggins #2 - Flight C337 -- (SBCFCD) 5 images
- 1981 Devil Canyon Basin - Flight C351 -- (SBCFCD) 3 images
- 1981 Devil Canyon Basin - Flight C390 -- (SBCFCD) 3 images
- 1981 East Badger Sycamore - Flight 419 -- (SBCFCD) 6 images
- 1981 Lemon Basin - Flight C393 -- (SBCFCD) 3 images
- 1981 North & West Badger - Flight C425 -- (SBCFCD) 4 images
- 1981 Twin Peaks - Flight 407 -- (SBCFCD) 6 images
- 1981 Twin Peaks Spreading Ground - Flight C408 -- (SBCFCD) 6 images
- 1981 Twin Peaks Spreading Ground - Flight C409 -- (SBCFCD) 6 images
- 1981 Twin Peaks Spreading Ground - Flight C410 -- (SBCFCD) 6 images
- 1981 Twin Peaks Spreading Ground - Flight C411 -- (SBCFCD) 6 images
- 1981 Twin Peaks Spreading Ground - Flight C412 -- (SBCFCD) 6 images
- 1981 Twin Peaks Spreading Ground - Flight C413 -- (SBCFCD) 6 images
- 1981 Waterman Basin - Flight C387 -- (SBCFCD) 2 images
- 1981 Waterman Basin - Flight C400 -- (SBCFCD) 1 images
- 1981 Waterman Basin - Flight C401 -- (SBCFCD) 2 images
- 1981 Waterman Basin - Flight C402 -- (SBCFCD) 2 images
- 1981 Waterman Basin - Flight C403 -- (SBCFCD) 2 images
- 1981 Waterman Basin - Flight C404 -- (SBCFCD) 2 images
- 1981 Waterman Basin - Flight C405 -- (SBCFCD) 2 images
- 1982 Mentone Dam Oblique - Flight C380 -- (SBCFCD) 3 images
- 1982 Scott Canyon Dam - Flight C424 -- (SBCFCD) 3 images
- 1983 Colton-Rialto - Flight C437 -- (SBCFCD) 64 images Index
- 1983 San Bernardino - Flight 75-83 -- (Imbiorski) 2 images
- 1984 San Bernardino - Flight C452 -- (SBCFCD) 1 image
- 1985 Harrison Basin - Flight C448 -- (SBCFCD) 9 images Index
- 1985 San Timoteo - Flight C448 -- (SBCFCD) 95 images Index
- 1986 Loma Linda - Flight 40-86 -- (Imbiorski) 2 images
- 1986 San Bernardino Valley - Flight C450 -- (SBCFCD) 184 images Index1 Index2 Index3
- 1986 Santa Ana River - Flight C451 -- (SBCFCD) 1 image
- 1987 Santa Ana River Wash - Flight C465 -- (SBCFCD) 8 images Index
- 1988 Santa Ana River - Flight C468 __ (SBCFCD) 22 images
- 1988 Santa Ana River - Flight C469 -- (SBCFCD) 9 images
- 1991 San Bernardino Valley - Flight C487 -- (SBCFCD) 215 images
- 1992 San Bernardino - Flight 19-92 -- (Imbiorski) 7 images
- 1992 CSUSB Devil's Canyon -- (Imbiorski) 1 image
- 1993 CSUSB - Flight 72-93 -- (Imbiorski) 14 images
- 1993 Temescal - Flight C501 -- (SBCFCD) 2 images
- 1994 Highland - Flight 6875 -- (NRCS) 34 images
- 1994 Redlands - Flight 6876 -- (NRCS) 2 images
- 1994 San Bernardino - Flight 6865 -- (NRCS) 43 images
- 1994 San Timoteo - Flight 6854 -- (NRCS) 8 images
- 1994 San Timoteo - Flight 6855 -- (NRCS) 2 images
- 1994 San Timoteo Canyon - Flight 6874 -- (NRCS) 183 images
- 1995 Santa Ana River - Flight C523 -- (SBCFCD) 89 images Index1 Index2
- 1996 San Bernardino Valley - Flight C528 -- (SBCFCD) 244 images Index1 Index2 Index3
- 1997 San Bernardino - Flight 76-97 -- (Imbiorski) 4 images
- 1997 Warm Creek - Flight C532 -- (SBCFCD) 29 images Index
- 1998 San Timoteo Creek - Flight C537 -- (SBCFCD) 7 images Index
- 1999 San Bernardino - Flight 37-99 -- (Imbiorski) 4 images
- 1999 San Bernardino - Flight 99-282 -- (Imbiorski) 2 images
- 1999 San Bernardino - Flight 99-828 -- (Imbiorski) 20 images
- 2001 San Bernardino - Flight 01-843 -- (Imbiorski) 10 images
- 2001 San Bernardino Valley - Flight C541 -- (SBCFCD) 254 images Index1 Index2 Index3 Index4 Index5
- 2001 Santa Ana River - Flight C542 -- (SBCFCD) 63 images Index
- 2003 San Bernardino - Flight 03-839 -- (Imbiorski) 6 images
- 2003 San Bernardino - Flight 03-843 -- (Imbiorski) 2 images
- 2004 Loma Linda - Flight 04-844 -- (Imbiorski) 3 images
- 2004 Loma Linda - Flight 04-845 -- (Imbiorski) 2 images
- 2004 Redlands - Flight 04-842 -- (Imbiorski) 4 images
- 2004 San Bernardino - Flight 04-847 -- (Imbiorski) 6 images
- 2004 San Bernardino - Flight 04-810 -- (Imbiorski) 18 images
- 2004 San Bernardino - Flight 04-809 -- (Imbiorski) 26 images
- 2004 San Timoteo - Flight C548 -- (SBCFCD) 12 images
- 2005 San Bernardino Valley - Flight C553 #1 -- (SBCFCD) 252 images IndexA IndexB
- 2005 San Bernardino Valley - Flight C553 #2 -- (SBCFCD) 252 images IndexA IndexB
- 2005 San Bernardino Valley - Flight C553 #3 -- (SBCFCD) 252 images IndexA IndexB
- 2005 San Bernardino Valley - Flight C553 #4 -- (SBCFCD) 176 images IndexA IndexB
- 2005 Upper Santa Ana - Flight C05-253 -- (SBCFCD) 82 images
- 2007 San Bernardino - Flight 08-802 -- (Imbiorski) 36 images
- 2008 San Bernardino - Flight 836 -- (Imbiorski) 27 images
-
Trona (178 images)
Cities that are included in this section include Trona
- 1930 Trona - Flight 1247 -- (Rowe) 153 images
- 1972 Trona - Flight C219 -- (SBCFCD) 6 images Index
- 1973 Trona / Argus - Flight C201 -- (SBCFCD) 18 images
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
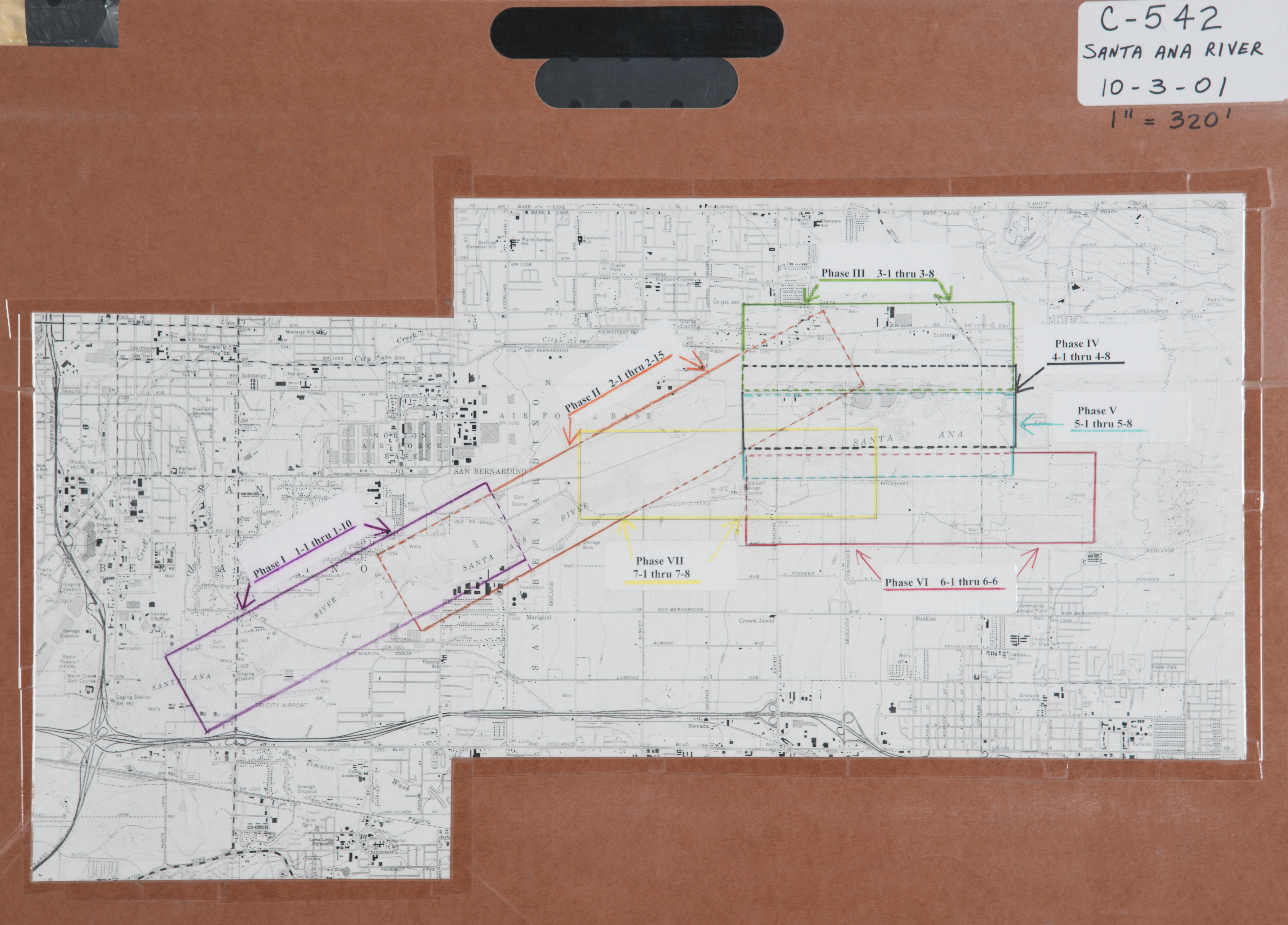{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
San Diego County (85 Images)
Cities that are included in this section include Valley Center and Oceanside.
- 1921 Pioneer-Misc. - Flight J23 -- (Rowe) 6 images
- 1921 San Diego - Flight J23 -- (Rowe) 5 images
- 1928 San Diego - Flight C300 -- (Rowe) 36 images
- 1931 Rincon - Flight C1740 -- (Rowe) 7 images
- 1939 San Luis Rey - Flight AXN271 -- (USDA-SBCFCD) 4 images
- Rincon Area - Flight 5405 -- (Rowe) 16 images
- Rincon Area - Flight 1740 -- (Rowe) 7 images
Santa Barbara (87 Images)
Cities that are included in this section includeGaviota, Santa Barbara, and Santa Ynez.
- 1929 Freeman Ranch - Flight 509 -- (Rowe) 4 images
- 1941 Freeman Ranch - Flight 7539 -- (Rowe) 7 images
- 1952 Freeman Ranch - Flight 18560 -- (Rowe) 28 images
- 1955 Freeman Ranch - Flight 22323 -- (Rowe) 16 images
- 1956 Point Arguelle-Santa Ynez - Flight GS-CDC -- (Rowe) 32 images
Ventura County (144 Images)
Cities that are included in this section include Ojai, Oxnard, and Ventura.
- 1928 Rancho Matilija - Flight 300 -- (Rowe) 1 image
- 1930 Rancho Matilija - Flight 883 -- (Rowe) 1 image
- 1938 Rancho Matilija - Flight 5029 -- (Rowe) 1 image
- 1941 Oxnard - Flight 7046 -- (Rowe) 1 image
- 1945 Oxnard - Flight C9800 -- (Rowe) 3 images
- 1945 Santa Clara - Flight 9800-5 -- (Rowe) 15 images
- 1945 Santa Clara - Flight 9800-7 -- (Rowe) 2 images
- 1945 Ventura - Flight C9800 -- (Rowe) 1 image
- 1947 Rancho Matilija - Flight 11730 -- (Rowe) 1 image
- 1950 Oxnard Area - Flight 14861 -- (Rowe) 89 images
- 1950 Oxnard - Flight 14861 -- (Rowe) 1 image
- 1951 Oxnard Area - Flight 16505 -- (Rowe) 12 images
- 1951 Rancho Matilija - Flight 16040 -- (Rowe) 1 image
- 1951 Rancho Matilija - Flight C18365 -- (Rowe) 5 images
- 1963 Ventura River - Flight G32 -- (Rowe) 4 images
- Ventura County - Flight 2902 -- (Rowe) 6 images
Yuma, Arizona (19 Images)
- 1949 Yuma - Flight 14613 -- (Rowe) 8
- 1957 Yuma - Flight 11400 -- (Rowe) 11 images
Imperial County (85 Images)
- Pre—1952 Imperial Valley -- (Rowe) 85 images
King County (44 Images)
- 1928 Kings River - Flight 348D -- (Rowe) 5 images
- 1928 Kings River - Flight 348E -- (Rowe) 4 images
- 1928 Kings River - Flight 348F -- (Rowe) 6 images
- 1928 Kings River - Flight 348G -- (Rowe) 5 images
- 1928 Kings River - Flight 348H -- (Rowe) 6 images
- 1928 Kings River - Flight 668 -- (Rowe) 18 images